ON CENTAURS
B. Lukács
President of the Matter Evolution Subcommittee of the Geonomy Scientific Committee of HAS
CRIP RMKI, H-1525 Bp. 114. Pf. 49, Budapest, Hungary
lukacs@rmki.kfki.hu
ABSTRACT
Classical Greek authors often report centaurs, hexapodal sophonts which roughly are man + horse compounds. Most reports are historical; but not all. While almost all modern authors agree that such beings never existed on Terra, it is interesting to see how not and why not. Namely it seems that a hexapodal land vertebrate body plan would have had advantages compared to the tetrapodal one. Then why Evolution did not lead to generalised centaurs?
0. ON LANGUAGE; OR EST HOMO MENSURA?
Xenophanes, the Eleata (even if he was originally a citizen of Kolophon; 580-484 BC) formulated an idea which is very probable even if we cannot check the hypothesis. He stated that if oxen, horses and lions could paint, then they would paint gods of bovine, equine and leonine forms. And a century later Protagoras of Abdera (481-411 BC) formulated the idea of "homo mensura" (of course, still in Greek; he did not speak Latin).
The "homo mensura" may or may not be good enough for a philosopher, but it is strange enough for a physicist. Einstein's ideal observers are not human; they have negligible mass, size &c., going to 0 in limit. Maybe a macaque of violet blue bottom could not be a non-idealised observer because his IQ is low; but any sophont ET can serve. With a very few exceptions, science's statements should be meaningful without any reference to humans.
Gorgias, the Sicilian (483-375 BC) for-+-mulated a rather provocative triadic statement. As far as we can reconstruct it from later references, the statement is something such:
1) There is nothing.
2) If something still exists by chance, that thing cannot be made known.
3) But if by any chance one could learn about that thing, the knowledge could not be given to anybody else.
It seems that he did not prove Statement 1. As for Statement 2), however, he gave good arguments about the ambiguities and individual differences of human senses. And for Statement 3) he listed several ways of misinterpretations in communication. These misinterpretations generally exist and act, except if we know about them and work hard to avoid them.
In our present topics Protagoras' maxim is very dangerous. We should use a quite nonhuman language, which is not easy and results in strange terms. Sometimes I will use strange and unstylish a language, indeed.
Let us see a few examples here. First, all Vertebrata have something which is straight in first approximation and is composed of vertebrae. This is called generally "the vertebral column". It is indeed a column in us.
It was such in Homo erectus and in the Australopithecinae, who very probably differed from us only on species level, but that point is irrelevant now. But in a dog, a salamander or a fish it is a vertebral rod. In a study connected with long-range evolution it is silly to speak about the vertebral column of an Eusthenopteron or an Ichthyostega. They did not have vertebral columns. The absurd language, of course, comes from History of Science. The anatomic language was defined during Rinascimento by humanists and physicians. They were quite far from any idea of biological evolution. From the beginning of XIXth century some biologists started to recognise the homology between the vertebral column of man and the vertebral rod of a dog; but then already the expression "column" was several centuries old. So they started to speak about the vertebral column of the dog, which is absurd. Observe that in a centaur the first half of the one-dimensional vertebral something would be a column, but the "posterior" half (not yet posterior, being exactly below the neck at the turn) would become a rod. In this study I write always rod. It was a rod for more than half of a Gy.
The second example for misinterpretation is communication is the number of "limbs". It is an interesting point, what is a limb. Maybe we could use the expression "extremities". Now, we always feel that there are 4 extremities, limbs or anything. A cat feels clearly that she has 5: four legs and 1 tail. I was able to observe some decade ago a cat which transiently lost the use of her tail, for a few weeks. She was completely baffled, and after some days she learnt to be very careful. She was, of course, able to jump up onto tables; but was not able to jump down, because her tail should have been working for equilibration. She was seriously crippled albeit less than without the use of a leg. A dog, being practically 2-dimensional, uses much less the tail. We do not have any tail, except a few rudimentary and small vertebrae; we do not see our tail, do not feel it, and generally do not have any experience about it at all; then it is a revelation to see a cat with an inactive tail. We do not even have any proper language to name the deficiency of the cat, meaning that we could not call the deficient cat "lame". Lameness would mean no full use of a leg, but not of the tail. Still, the cat was crippled.
Now, there is no doubt that present land vertebrates originate from "fishes", in the sense that originally their ancestors lived in water, and they would have been classified amongst "Pisces" (and they, therefore, are even then in that time no sophont observer was present). Interestingly enough, the idea goes back to Anaximander of Miletus (611-546 BC), who observed the great maternal care needed for the human baby while not for an immature fish.
Now, the "body plan of fishes" generally included 8 extemities: 2 pairs of paired ventral fins, one anal ventral one, two dorsal ones and one caudal one. A few million years before the transition Eusthenopteron still had all the 8. 3 are lost in any relatives on land, and 4 are lost in us. We cannot easily imagine us as fishes.
When we try, we have the tendency to call the anterior pair pectoral, and the posterior pair pelvic. And the absurdity is not obvious in English, but it is in Latin, German or Magyar.
The Latin "pelvis, -is" is "dish" or rather "tureen"; or "basin". The secondary meaning is the English "pelvis". It is easy to understand how the anatomical meaning started: the complicated assembly of 3 bones, the ilium, ischium & pubis, form in us something mimicking a deep tureen or a basin. In German in both meanings the word is das Becken; and also in Magyar: "medence".
However this compound did not exist before the first really amphibious Crossopterigii. In a fully aquatic lifestyle the bones of the posterior paired fins were not needed to be attached to the vertebral rod; and indeed they were not attached. So nothing tureen-like compound existed about the stem of the posterior paired fins: no pelvis. Still people call something at the stem pelvis; even pelvic girdle. This may sound as being simply finicky; but it is not. Namely, Eusthenopteron has two fins attached to the vertebral rod; but they were not the posterior paired ones, so the "pelvic" ones in the anthropomorphic language. (See in due course.) This will be the reason calling here the posterior paired fins not pelvic but abdominal.
And so on. Sometimes in the main text I shall repeat statements mentioned here.
1. INTRODUCTION
Ancient Greek tradition had stories about centaurs: people with four legs and a human upper half. It is difficult to tell the primitive form of the tradition and it is doubtful if the Mycenian age had anything about them; but Herakles, prince of Mycenian-age Thebes, fights them, and they are depicted in the Hero's Tomb in Lefkadi, which is just after the collapse of the Mycenian civilisation. Homer mentions some "pheres" (obviously Classical "theres"=animals, beasts &c.), but without details, although one of his stories is the Classical Centauromachia, the centaur-lapith fight at the feast of King Pirithous & Queen Hippodameia. (Hippo- is horse-!)
Later, in Classical ages the stories stabilize. Similarly, beauty ideals for centaurs as well [1]. Centaurs have equine limbs, tail and rump, but it continues in a human upper half. So a centaur has a tail, four legs and two arms. But it is obviously a mammal, and sometimes the males rape human females. (E.g. the Centaurs at the wedding of Pirithous and Hippodameia wanted to rape Hippodameia, which is not contrary to common sense.) We do not hear about centaur females for a time, but later they are also mentioned or depicted. Centaurs are sophont, and Kheiron the Centaur was wise, e.g. he was for a time the tutor of both Achilleus and Asklepios, but generally the centaurs were wild. They definitely did not tolerate wine, both in the sense that they committed anything to get it, and also that then they became uncontrollable. The famous Lapith-Centaur battle at Larissa broke out because they got some wine and then wanted to rape the Queen.
According to common opinion, the first centaur was the product of the copulation of Ixion & Nephele, the latter being the simulacrum of Hera. Ixion wanted to seduce Hera and Zeus wanted to make him monkey [2], [3]. Then, maybe, Centaurus copulated with the mares of Magnesia [4]. (Magnesia was horse country.)
What is interesting: Greeks of Classical ages did believe in the existence of centaurs. OK, they were more frequent in earlier, simpler times (just as satyrs, nymphs &c.), but...
Plutarch in the Feast of the Seven Sages [5] describes a meeting of the Seven Sages at the court of one of them, Periandros, the Tyrant of Corinth. The sages would just go to eat, but horse shepherds bring a newborn centaur for show. Some Sages are afraid because of the hidden meaning. Then Sage Thales tells that the horse shepherds of the Tyrant behave themselves badly; and then the Sages go to feast. While this story may be a satire, Pliny tells that he saw the body of a Centaur, preserved in honey [6]. The story is continued under Emperor Hadrian by Phlegon of Tralles [7]; he also saw the centaur preserved in honey. From the two authors we learn that during Emperor Claudius somebody discovered a herd of Centaurs "in a mountainous region of Arabia", at a place called Saune. One of them was captured, for being sent to the Emperor via Egypt. Unfortunately, he died in Egypt, but the Praefectus Aegypti ordered him to be conserved in honey, and the Emperors kept the gift at least until Hadrian. On the other hand Lucretius Carus does not regard the centaurs viable, but the argumentation is restricted to the quite different maturation ages of the human and equine parts [8].
So, in classical times Centaurs were accepted by the majority but they were believed to live in not too populated hilly regions in Magnesia, Thessaly and Arcady. Only the extremely sceptic few (and the scientists; I mean hard science, see 3 examples in Appendix A) did not believe in them.
Centaurs are quite harmonious creatures as one can see from sculptures; otherwise nobody would have believed them at lest for one millennium and half. Lucretius' counterargument was not bad but was not too strong and anyways ineffective. And there are "stranger" (I mean, more different from the "usual" groups of animals) body plans represented in the Cambrian Burgess Shale than a mere hexapod using the frontal pair of appendages as manipulators. (Generally a phylum means a unique body plan. The number of extant phyla is 2-3 dozen. As for the extinct ones, who knows?) Lots of strange fossils can be found in the Burgess Shale.
Opabinia regalis is a quite good species, with few individuals, more or less complete, with maximal length 7 cm. The body seems segmented. However the head has 5 (!) eyes, the mouth is turned backward, and in addition from the top of the head starts a proboscis. Of course, one may reconstruct a way of feeding via the proboscis, still it is strange that the mouth is in a place unfit for direct feeding. Now, there are two serious suggestions:
1) that the animal was a not too close relative of both arthropods and onyxophorans, from a now extinct phylum [9]; or
2) that the phylum is extinct even with its relatives [10], in which case it is rather difficult to guess its place on the evolutionary tree.
I emphasize that more or less each phylum means its own body plan; so indeed Opabinia regalis is
more unusual than Centaurus sapiens; still it existed.
The second surprising Burgess Shale animal is Hallucigenia sparsa. Its length is not bigger than 3 cm. It seems that it has 7 segments, maybe a head in the front (there the preservation is wrong), a tail or body end on the other side, and, according to the first interpretation [11], 7 pairs of legs below, 7 unpaired tentacles on the dorsal side in the medial line, and behind, but before the anus or anything, a few short tentacles. The idea was that the dorsal tentacles mediated some food to the mouth. The taxonomy was again rather doubtful. Of course, Gould's opinion was an extinct phylum. Then a related species was found in Chinese Cambrian layers and a new reconstruction changes the dorsal direction to ventral and vice versa, and reinterprets Hallucigenia as an onyxophoran. However it is far from sure, and again, H. sparsa and H. fortis seem stranger than a centaur.
Finally, I mention the Late Precambrian Ediacara fauna (?). One can find almost any interpretation of them. Some experts are not even sure that they were Animalia; it may be an extinct Kingdom. Others interpret them as animals but claim that some Ediacarans did not have any mouth (!), so these individuals made their intake and void via diffusion. Others claim that the fossils are Cnidaria, Arthropoda &c., but rather strange ones. For a new Kingdom Vendobionta see [12]. For a quite unusual Ediacaran body plan see [13]. For later discussions see e.g. [14]. For considerations about symmetry & topology see e.g. [15].
Now, if such animals did live, why just centaurs did not? Are hexapod Vertebrata impossible if Hallucigenia, Opabinia and mouthless Ediacarans are not? And why and how?
2. FOUR LEGS GOOD, TWO LEGS BAD
Of course, this slogan is from Orwell's Animal Farm. However, it is as well a general mammal experience, shared by Reptilia & Amphibia (but not Aves; the phylogenetic causes are coming, in due course). The reason behind is that for stable support you need at least 3 points; but then still you may topple when walking and one leg does not touch the surface. So the minimal safe number of legs is four. It seems that the maximal safe mass even then is in the order of 105 g [16]; this is true for land animals on Earth, but maybe more or less for the whole Universe, see App. B. Also, there were interesting counterexamples in the geologic past (and maybe even present), which is disturbing; see also App. B.
Humans, however, walk erect, on 2 legs, because the forward 2 is used on manipulation. (The liberation of a pair of legs from locomotion is necessary for using tools which is then necessary to trigger development of intelligence. It is rather doubtful if an animal could develop intelligence without free hands. Dolphins may be a borderline case.) After 4 million years of experience they can walk on only 2 quite safely except on icy terrains; but that they are at the safety border for mass is shown by the fact that in advanced age, when bones become more fragile, some femoral fractures, lethal even now in some 15%, do happen. However, this happening in postreproductive age, it is not a barrier for mass. See also the serious mass increase between Australopithecus/Homo africanus and Homo erectus, parallel with the improving bipedalism. Birds also walk on 2 legs, the frontal ones having turned into wings, but the flying ones are light, and Ratitae can use the wings for equilibration.
However, obviously, 6 legs would be even better than 4. Several arguments are possible. I mention here only 2.
1) Lots of mammals move in a way that in one moment only 2 legs touch the surface: e.g. fore left & hind right, while fore right & hind left step forward, and then they stand and the other 2 step. Sometimes it is told that this pattern goes back to the swimming of aqueous Crossopterygii [17]. Such mammals can topple; and leg fractures are frequent for horses, after which they frequently cannot recover. Of course, pre-domestication horses were smaller; and also, a tetrapod generally can stop the motion when starting topple; but not always. With 6 legs they would be stable even if stepping simultaneously with fore left, middle right & hind left.
2) With 6 legs the intellectual evolution would have been faster. Namely, the 2 forelegs could have been manipulating well back in evolution. See the discussion in P. Anderson's Fire Time.
No surprise that some sci-fi authors expect hexapodalism to be the leading forms on lots of alien planets; here I mention only 2 such distinct authors as J. Pournelle in Starswarm and the Tran trilogy and F. Carsac in Les Robinsons du Cosmos. Also, P. Anderson (who graduated just as a physicist) used the hexapod theme not only in Fire Time, but also for an unrelated sophont species on an unrelated planet of high gravity in the van Rijn novels and in the Flandry novel Game of Empire. The recent successful 3D movie Avatar has a lot of hexapod animals too. And still, we are tetrapods...
We more or less would expect that Evolution be a competition via Struggle for Life. But then we would expect a hexapod variation of Vertebrata. Even if 6 legs were not present at start (and we shall see later that the question is not quite trivial), it might have appeared as a rare mutation and then the new form may have passed the tetrapods. There was more than 350 My for this. Why not?
3. ARE WE REALLY TETRAPODS?
Fishes, amphibia, reptiles, birds & mammals are grades, not clades. The descent is shown by cladistics. Before Devonian all the leading Deuterostomata were "fishes", in the sense that they lived in water. The land was almost empty, populated mainly by moss and small arthropods. However, one group of "fishes", roughly Crossopterygii and maybe lungfishes, started something interesting. Modern systems generally call the "progressive" clade Choanata (generally living lungfishes are not counted amongst Choanata); some Choanata have fins, some legs. According to older understandings of Evolution (Late Nineteen, Early Twenty centuries), the legged ones were/are the land animals, and they are the Tetrapoda. Surely, according to elementary Greek, only legged ones are proper to be called Tetrapoda; still the transition from "fins" to "legs" is rather continuous when you have enough fossils. I will use here the cladogram of Coates [18]; of course, new fossils may rearrange the proper cladogram.
The question in the Chapter title has at least two disjoint aspects: 1) 4 or not; and 2) legs or something other? I handle these 2 separately.
4. FOUR OR MORE?
It is generally believed that the number of "extremities" of "higher Vertebrata" is exactly four. While this is true from "Amphibia" for legs, observe a cat, whose tail is unable to move because of some accident. I was able to follow the process of recuperation of a feral cat, together with colleague Ágnes Holba, for weeks. The cat had been hurt by a broken branch of a tree at the stem of her tail, first the tail could not have been moved, later the use started to come back. When the tail was out of use, the cat was very much restricted in propagation, so she was "lame". Namely, cats propagate vertically too. But without a movable tail the cat could not get down from places where she of course could get up without problem. So, while legs and tail differ in Tetrapoda, there is something common among them as well. Humans, having practically no tails for at least 20 My, are not the proper classifiers in caudal matters. Of course, tetrapod tails are not quite homomorphic with the legs, because the tail does contain chorda dorsalis & vertebrae while legs do not; but even this is not so simple.
Life started, for obvious and inevitable physical & chemical reasons, in continuous fluid phase, be it a pond, lake, sea or anything. Being the physical laws the same everywhere, this is true even for extraterrestrial life. On Earth intelligence developed on land, so the transition from almost weightless situations to full weight was critical and very important for the number of legs.
We have at least one living fossil, Latimeria chalumnae, which is a good Crossopteryx. In it, the four fins homologous to tetrapod legs are robust and contain bones (even if not in contact with the vertebral rod (which is a column only in humans, of course). However it has dorsal fins as well, and the dorsal fins had bones even in Cretan Macropoma [17]; and of course, the caudal fin has the end-vertebrae within. Also, Latimeria has a ventral fin behind the paired "hind-fins" as well, and the same was true as well for Middle Devonian Eusthenopteron [19]. Note that while Latimeria, Macropoma and Eusthenopteron are all Crossopterygii, they represent at least 2 trends: the first 2 did not show tendency to become land animals. They may or may not belong to the same clade; but Coates' cladogram [18] indeed constructs a branching away sequence:
Coelacanths (or Actinistia) - lungfishes - Eusthenopteron
Eusthenopteron is anyways in the clade leading towards Tetrapoda; but it still has dorsal fins as well as an extra ventral one.
Panderychtis, branching away subsequently, seems to have lost its fore dorsal fin (remember this); maybe the hind dorsal and the extra ventral ones have been incorporated into the enlargened caudal one, maybe they were lost. So Panderychtis has only 5 extremities, as cats: two pairs of ventral fins, and a caudal fin.
Interestingly enough it seems that the first Vertebrata had no paired fins at all. Amphioxus, which is not a Vertebrata, but a Cephalochordata, but which is near enough, still has only one medial fin traversing most of the body first ventrally, then caudally and finally dorsally. Agnatha are clearly Vertebrata (lamprey), but extant Agnatha also do not have paired fins. So far the first fishes with paired fins were the Silurian Acanthodii. Maybe first there was a paired long fin, and then they separated into pairs of fins [17]; but some Acanthodii had as much as eight pairs. Then appear the proto-sharks and they have exactly 2 pairs. Maybe simply 2 pairs at the best places for fast & precise swimming. Who knows?
5. FINS OR LEGS? LEGS OR ARMS?
It seems that Crossopteryx paired fins are homologous & homomorphous to tetrapod legs, so it is more or less a matter of preference when we call them fins and when legs. Also, we may call them fins on aquatic animals and legs on land ones. Or we may call them legs when already the extremity ends in the "palm", 5-digited, of course. But there are problems with all these views, except of course the indecisive first.
Look at extant amphibians. Coecilians are definitely land animals; frogs are more or less land animals when adults, but extremely aquatic tadpoles when larvae. As for Urodela, almost all of them like the half-aquatic environment and the axolotl is purely aquatic. As for extinct amphibians, in many cases we cannot tell, in which extent the animal was aquatic (no surprise; they were amphibian). So at which point(s) of the evolutional tree did they become land animals?
And as for 5-digit palms, Acanthostega had 8-digited legs, Ichtyostega was 7-digited and Tulerpeton 6-digited [20]. Even more disturbing is the case of Ichthyosauria. Following Huehne [21], the order lived from Lower Perm to Uppermost Cretaceous, and became extinct with latipinnate Myopterygius. During this we never see a palm-plan, and the animals seem fully aquatic. This may be an early adaptation of reptiles to secondary aquatic life (as for whales amongst mammals; the whole Carboniferous was long enough for this), but also it is possible that the precursors of ichthyosaurs remained aquatic and never gave up fins. Huehne prefers the second scenario. Now Ichtyosauria are generally classified as reptiles/amniotes; still they have fins.
The second problem can be first approached from Primates viewpoint. Sometimes Primates are regarded as having 4 hands. Namely, monkeys & apes can catch with all of four palms. Then the homologous arms & legs can even be understood as four arms, not four legs. Of course, we have the tendency to appropriate legs to mere animals, not arms; but this is superstition/chauvinism.
Now, it is possible that the first leg-like extremity was rather arm not leg. As an analogue, consider extant Exocoetus volitans, in vulgar name the trigla or “swallowfish”. It is a rather archaic teleost, so it is not in our clade. Still, the pectoral fins have 3-3 digits. The trigla can walk on them; but the hind paired fins did not form digits. (See App. C about the trigla.)
Indeed, it would be easier to form real well-developed arms than legs for "fishes". Namely the pectoral fins are just behind the bony skull, so the bony supports of the arms could immediately join to the base of the skull, while for the legbones of the hind fins there is no such place for a joint in any extant "fish". I will return to this point in due course.
For "common sense" amniote reptiles are par excellence land animals, so their legs must support the full weight of the body, so they must have well-developed legs, contrary to many amphibians, half-aquatic, and the weight is effectively cca. one-tenth in water. Now, on Coates' cladogram Amniota branched earlier (c. 365 Mya) than Amphibia (c. 340 Mya); the sister taxon to the first one is 6-digited Tulerpeton, while to the second it is Greerpeton, and Greerpeton's hindlegs seem more active and more terrestrial than that of Tulerpeton. But this is something the "common sense" learnt from extant amphibians & reptiles. Neither "Amphibia" nor "Reptilia" are true clades but grades. Amphibians are the recent amphibian vertebrates, so those which did not completely leave the aqueous environment, while reptiles are such recent vertebrates which have left it (except for some turtles). Neither group is necessarily monophyletic. Indeed, several authors tried with a diphyletic Class Amphibia and a polyphyletic Class Reptilia; the question deserves an Appendix D. Of course, the cladistically defined Amphibia must be monophyletic; then the question is exactly which animals belong thither.
6. (HIND)LEGS BECOME ATTACHED
Various scenarios have been suggested for the process in which some Vertebrata became land animals. The oldest scenario was the great droughts of the Devonian (the Age of the Old Red Sandstone): separate ponds had been formed and if one started to dry out, the most adventurous lungfishes/lobefins got out and slowly crawled to the next one. (Without the uplift of water they were unable to properly walk even if they had lobe-fins of even already hands.) Of course the stronger support gave greater chance for survival, so Struggle for Life/Evolution led to stronger legs. Good; but for support without uplift the legbones must be attached to the vertebrae. OK; they became attached. However, that is easy and immediate for the bones of the pectoral fins: they can be attached at the base of the skull. But for the hind fins it is not trivial; for two reasons:
1) Whither to attach? The row of vertebrae in the hind parts is/was almost translation-symmetric, no preferred point. (The sacral region comes later. The vertebral rods of extant fishes do not show inhomogeneities in regions guessed to be homologous to sacral regions of amhibians, reptiles &c., so it seems that the sacral region is not predestined.) So there was no "natural" place to attach, and this may hindered the process.
2) Even if already the legbone touches the row of vertebrae, a proper joint must be formed, simple fusion would yield support but not motion. This is a complicated process.
These 2 points would suggest that the transition to (land) tetrapods happened exactly once. However Huehne [21] prefers 2 independent transitions: one to Urodelidia and one to Eutetrapoda. (The idea originateds from Säve-Sôderbergh and then from Jarvik; see Appendix D.) He observes that in Urodelidia the forelegs appear earlier in ontogeny than the hindlegs and in many cases the forelegs are the stronger; while it is oppositely in Eutetrapodes.
This view was supported by evolutionary trees where the Eutetrapoda originated from the Crossopteryx order Osteolepiformes, while the Urodelidia from the neighbouring order Porolepiformes (see e.g. [22]); the argument for the latter was embryologic. The argument was weakened by the narrowness of the paired fins of the Porolepiformes.
Anything may be true, but for any case nobody suggested more than 2 transitions.
Closely related Ichthyostega & Acanthostega are the oldest known tetrapods with expressed pelvic joint and a neck region. (Ichthyostega a little bit younger and much more documented. Tiktaalik a little older with neck but we do not yet know about its pelvis. Eusthenopteron had something called a pelvis and something other similar to a pelvis; but no neck.) But it seems even they lived mainly in water. So even the very powerful pelvic joint does not seem to have been enough for support without uplift.
The probable explanation is that a long straight horizontal row of vertebrae cannot work on land, because in the middle it would sag: an arch is needed. For this, however, the vertebrae must be wedged. The initial, unwedged structure is the embolomeric, while the final structure is either the stereospondylic or the gastrocentral; these two are rather equivalent for efficiency. The transitional structure is the rachitomic.
You cannot have a multiplicity of legs because originally you had only a few fins. (True, more than 2 pairs. E.g. the recent coelacanth, the Latimeria, has a third, dorsal-ventral, pair, behind the homologues of our hindlegs as well.) So you must form the arch between the supports. Until a good arch is not ready, you need the uplift. (On the other hand, forming the arch is much faster than forming new legs ex nihilo.)
Of course the par excellence aquatic lobe-fins had embolomeric vertebra. For Ichthyostega it was still embolomeric [21]. Maybe the strong pelvis was useful for relatively fast motion; but you cannot be at home on land with embolometric vertebra.
So either Acanthostega & Ichthyostega remained mostly in water because the vertebral row was still embolomeric, or it remained embolomeric because they did not yet leave the water. Then the more terrestrial descendents wedged out their vertebrae more and more and got nice arches. Lower Permian Cacops is already rachitomic and the whole order stops at this stage.
7. VERTEBRATA AS COLONIES; OR WHY NOT A MULTITUDE OF LEGS?
At the end of the XIXth century Herbert Spencer defined primary, secondary and tertiary animals (see e.g. [23]; in this point his language was somewhat clumsy). Definitions are as follows. A primary animal cannot be divided into living parts; so they are unicellular. A secondary animal consists of primary ones, i.e. cells. A tertiary animal consists of secondary ones, e.g. the Arthropoda or Annelida have repetitive parts which originally were very probably secondary animals. His opinion was that Vertebrata are secondary animals; the repetitive characters are simply adaptations to, e.g., swimming. While this last statement seems forced, let us first note that now the steps must continue both down and up. Namely, the Serial Endosymbiotic Theory of Lynn Margulis suggests that Eukaryotes were formed in endosymbiotic steps and the first one was the endosymbiosis of a termophile Archaea, and a few Spirocheta-like eubacteriae, maybe near to recent Hollandina [24]. If so, then the primary organs of motion (flagella, undulipodia) were "at zero degree" independent predator bacteria. While this 0th step of integration is not (yet?) fully accepted, it may quite be true. See e.g. [25] and citations therein. On the other hand, Marais suggested [26] that the termite colony is a higher organism composed of termites. Because a termite is tertiary, the colony must be quaternary.
Now let us go back to proto-Vertebrata. Surely it was Precambrian; but it must have been near to the ancestors of extant Amphioxus (of which Pikaia is found in the Cambrian Burgess Shale), and it must have had some relation to the ancestors of present Tunicata as well. Now, neither Amphioxus, nor Pikaia, nor any Tunicata have legs. Interestingly enough, the Vertebrate Archetype reconstructed by Owen [27] in definitely non-Darwinian ways is rather similar to Amphioxus.
Now, Amphioxus is clearly segmented. It has dozens of gills, pulsating arteries (instead of hearts), protonephridia and "eyes" (sensitive cells) in segmental arrangement; and while less gonads than the gills, still at least 2 dozen pairs, again segmentally ([28], [29] and citations therein). It is highly improbable that adaptation to swimming form eyes & gonads segmentally repeated. But then there may have been a possibility of original multiplicity of legs, of which either 4 or 6 may have been enhanced.
However this cannot have happened. First, because very probably segmental legs (well-known on, say, insects) never existed on proto-Vertebrata, or at least were lost well before Devonian. Recent Amphioxus has no legs or any appendages except "tentacles" around the mouth and a continuous medial dorsal fin. Definitely no paired appendages are present [28], [29]. Recent Agnatha also have only unpaired fins, most probably remnants of a fin continuous both on the dorsal and on the ventral surface in the symmetry plane. Owen's Vertebrate Archetype did not have appendages as well. So we may conclude that proto-vertebrates did not have legs. Then the few fins are innovations for e.g. swimming, and we cannot expect many legs at the Devonian startpoint of walking.
As for virtual legs; in very recent years we might know and tell something more or less constructive, because of the Hox families. See Chap. 9, the Conclusions, and for the details, Appendix E.
8. AND THE HEXAPODA INSECTS?
Interestingly enough, insects do fulfil the Hexapod Body Plan, characteristic for centaurs and for the products of many sci-fi writers. In first approximation, insects have always 6 legs.
However, observe 2 serious differences between insects and centaurs.
1) Insects are rather small. Indeed the tracheal way of oxygen intake is a fundamental restriction for insect sizes. So no supportive skeletal row is needed, even on land; and
2) The 6 legs is the endpoint of evolution from a >>1 initial state. This Point 2) deserves some further discussion. Here I do not give References. The insect literature is rather tremendous, and I think I could find support for any statement which I can imagine. Still, what I am going to tell is rather elementary.
All insects originally had 21 segments. (Surely, because they are monophyletic.) Sometime in the far past first the originally rather homogeneous repetitive pattern differentiated into 3 regions: head (6 segments), thorax (3 segments) and abdomen (12 segments).
Now, the thorax has 2 walking legs on each segment; that is 6 altogether. However originally the abdomen had legs as well, and even now at least some segments of the abdomen may have rudimentary ones. E.g. the Protura (e.g. Eosentomon transitorium) have rudimentary legs on the abdominal segments 1, 2 & 3. The thoracic segments of the Apterygota are undifferentiated, showing the archaicity of the group; and indeed there are rudimentary legs on the abdomen. The Diplura (e.g. Campodea staphylinus) have a rudimentary legpair on the abdominal segment 1, styli on segments 2-7 and a pair of cerci at the end of the abdomen. The Thysanura (e.g. Lepisma saccharina) do not have par excellence legs on the abdomen, but they too have styli & cerci.
As for the head, the organs around the mouth, the mandibula, maxilla & labium, are derived from the appendages of segments 4, 5 & 6. Segment 1 bears the paired feeler. So only the appendages of segments 2 & 3 have vanished.
Since the supposed relatives of the insects have a tendency to have a pair of legs on many segments, we may as well stop here.
9. FROM RUINS INTO A NEW WORLD
There is still argumentation why the "progressive Crossopterygii" went to the land. However now we know, as e.g. Aristotle did not quite recognised [30], that why has a double meaning: because of what and for what. Obviously a number of purposes can be listed. E.g. if Late Devonian had been extremely dry, a general view in the first half of last century, then a fish could have found useful to develop strong and sturdy fins to traverse dry spots between a drying up and a watery pond. But also, lands were a new world for vertebrates, populated only by plants & insects, and without competition for food. And so on.
Ref. [31] argues for a thermal purpose. If a fish can go to land for basking in the sunshine, its blood heats up, its activity speeds up. Then, returning to the water, it can catch more fish. By other words, the most progressive lobefins of Late Devonian were the top predators of waters; they, as Antaeus, from time to time they went out of water to touch Mother Gaia, the Earth, accumulated the energy from Helios, and went back to feed again.
OK; but Ichthyostega was not a top swimmer. I think it swam quite well; but not so well as the prey. Namely, it had only one remaining fin, the caudal one. His uncles, Panderichthys & Eusthenopteron possibly were more versatile in the water.
To see this, let us count fins on "fishes", meaning Pisces. The general plan is quite constant; of course an individual group may lose some fins. Going from the end of the skull posteriorward we find them as follows.
The pectoral fins, paired.
The abdominal fins, paired and one dorsal, cca. in the same position along the axis.
The anal fin ventrally and again one dorsally, again cca. in the same position along the axis.
The caudal fin, at the posterior end.
This is four positions along the axis. An educated guess is that ventral and dorsal fins have similar places along the axis because it is parsimonious for the Hox genes to organise the axis so. The 4 positions for 8 fins are obvious on the relict coelacanth Latimeria, and it was so until Eusthenopteron in Late Devonian. But observe that its older contemporary Panderichthys, possibly nearer to the survining lineage had only 5 fins: the homologues of tetrapod limbs & tail. Still, they were true fins.
Experts of hydrodynamics and builders of gliders could tell us why one or other plan is better to swift or versatile swimming. Some fishes use their fins for active swimming, some use only the caudal fin for propulsion, and use the others as rudders; & so on. Still, I would be surprised if Panderichthys could not have evaded Acanthosthega, had they been contemporaries (they were not) and if Eusthenopteron would have not been better in water than Panderichthys. Both had fins where Acanthostega & Ichthyostega had digits; and Eusthenopteron had 8 fins instead of Panderichthys' 5.
Well, Eusthenopteron seems to have been adapted to crawl out of water, in the sense of [31]: to crawl just out, for basking. It had a pectoral girdle, firmly supported by the skull. The abdominal fins were not attached to the vertebral rod; but the anal one was! (See [31], or, for more details, [32].) Well, the opposite dorsal one was attached too, and surely the dorsal surface was not used in the crawl; but maybe the Hoxes & other genes worked in such a way. (Hox genes, as we shall see, organize the body parts of a "linear" animal anterior-posteriorly; in Drosophila along the 21 segments, in Vertebrata along a few dozen somites/vertebraes &c. I return to the problem in next Chapter.)
Now, why to have a bony connection for the anal fin, if the abdominal ones do not have it? Maybe the bony connection helped to support marginally the body and move it on land. Only its own length should have been traversed; and note that the foremost and hindmost ventral ones had the bony contacts. After a fashion Eusthenopteron had “pelves” too; but they hand no bony contact with the vertebrae.
So you can imagine Eusthenopteron first crawling out of water slowly; then basking in the sun; and then going back at once and swimming rapidly and dexterously "as the lightning" (albeit this simile is somewhat out of place underwater).
And then the Firmament collapsed onto the basking Crossopterigii. Almost literally so.
Late Devonian is a well-known "global extinction" period, as e.g. the C-T and P-Tr boundaries. The causes are, of course, more and more difficult to find as we are going pastward. The "dinosaur killer" was almost certainly an asteroid, but the initiator of the (much more extensive) P-Tr extinction is not found yet. However it seems that the Late Devonian extinction coincided with an impact, so very probably the impact triggered it. Impact craters from that time are known, and at various localities, quite dispersed, spherulae can be collected as well. See e.g. [33], [34], [35], [36]. E.g. [33] mentions the Siljan Ring, Charlevoix Crater, Senzeilles & Qidong spherules, as consequences of the impact. While spherulae can be produced in other ways than impact too, they at least corroborate the other signals of impacts. Charlevoix Crater is some 50 km in diameter so the impactor was at least 8.
Big impacts disturb the biosphere transiently; still some consequences may persist. By any chance the Dinosaur Killer (9 km) caused only a few years of "nuclear winter"; still it has terminated lots of lineages. Not only dinosaurs; ichthyosaurs became extinct too, and the enantiornithes, the "other birds", dominating then the Northern Hemisphere. Also such successful protostomians as the ammonites. We do not fully understand, why.
Now, spherule collection helps to sharpen the dating of the event, and it seems it did not happoen at the D-C boundary, but within Late Devonian, at the Frasnian-Famennian boundary [33], [34], [35]. And lo; Eusthenopteron is Fraasnian, while Acanthostega & Ichthyostega are Famennian. Evolution of the "first tetrapods" was as fast as that of the great apes (including us). Let us see a few numbers of absolute dating even if changes may come in the future (lower boundaries, Mya):
|
Frasnian |
(U. Devon.) |
377.4 |
|
Famennian |
(U. Devon.) |
367 |
|
Hastarian |
(Tournaisian, L. Carbon.) |
362.5 |
|
Ivorian |
(Tournaisian, L. Carbon.) |
353.8 |
|
Cadian |
(Visean, L. Carbon.) |
349.5 |
So when the purging tempest has passed, no progressive Crossopteryx is seen anymore; there are such ones which do not want to see the lands, and a few descendants of the deceased, with digits instead of fans of fins. They are tetrapods. They completely lost the dorsal and anal fins, so there remained nothing, even buds, for more legs. This may have been the moment to loose Hox-14's (see App. E).
Maybe the catastrophe at the Frasnian-Famennian border made the transition to tetrapods so rapid. After the catastrophe there remained only 4.5 My from the Devonian; humans exist only for 5-5.5 My.
For tetrapods all the vertebral rod after the sacral region is caudal. But the Hoxes must have somehow marked out the position of the anal & opposite dorsal fins; we see on Eusthenopteron that they were bonily attached to the vertebrae. This region is completely absent in us [37]; it is definitely not the sacral one, which should be at the line of the abdominal fins. It is still rather difficult to understand how it might have vanished if the defining Hox remained.
Of course, an anal ventral fin is no good if you crawl. Maybe it was better to get rid of the most posterior ventral appendage, indeed. Maybe in a long enough time the anal-dorsal pair may have been transformed into a third ventral pair; but there was no time. The Frasnian-Famennian catastrophe disturbed the slow, gradual evolution.
The present structure is not unknown even before the catastrophe, however. First, Panderichthys, without anal & dorsal fins, is Frasnian. Second, Alberg reports an isolated Frasnian bone, which may be a pelvis [38]. He believes the bone belonging to i) a pelvic girdle of ii) a tetrapod from iii) the Elginerpeton genus. Then the tempest purged all beings going to land with 8 fins. Who knows why?
10. CONCLUSIONS AND OUTLOOK
First let us very briefly pass the (im)possibility of hybrids. Humans and horses differ on higher than generic level. Viable hybridisation does not have a chance, gene actions are incompatible (this is the modernised variety of the old counterargument of Lucretius Carus), and the common mammal body plan would result in tetrapod offspring even if… (human & horse hindlegs are homologous, so they would not add up). If one believes that I do not know about the very surprising transphylan fertilisation reports, read Appendix F. I know about them, I am not sure about them, but they are irrelevant in centaur context.
However, I think Hox genes show us something about a lost chance for higher fate for land vertebrates.
Hox genes are sets of genes which help to organise bilateral (segmented?) animals' bodies. I will not go into genetic details, because geneticist I am not, and the whole area of Hoxes is still in rapid development & change; and even the majority which will be told, went into Appendix E. However even without the very details the topic is important and interesting. As we will see, Hoxology may tell us sometime about the formation of first legs, definitely "why 4?".
Let us go back first to Classical Philosophy. As mentioned in Chap. 7, Herbert Spencer distinguished 3 different "levels of organisation" in animals. Now consider an animal of 3 levels: cells, repeated "segments" composed of cells and the animal, the whole, composed of the segments. An obvious example is the annelids, composed of many almost identical segments, "rings"; but the tertiary structure is clearly recognisable also in Arthropoda, Myriapoda &c. Spencer was sure that Vertebrata are not tertiary, the segments are only secondary adaptations for swimming or such. However see such contemporaries of Spencer as Hatschek [28] and Sedwick [39] (and of course see me now [40]). Amphioxus was generally regarded a segmented animal; and Vertebrata are related.
Now, XXth century's political life would have been less tragic if an analogy had been clearly recognised: sometimes Unification is not good if forced to extremes. In recent European Union the idea is called subsidiarity and believed to have originated in Christian Democratic political philosophy; but in 1991 almost anybody in almost any country believed in the Unified Southern Slavic State, except the Southern Slavs themselves; and believed in One and Indivisible Czechoslovakia, except for the majorities of Bohemians & Slovakians. XXth century's Philosophy was for as big unified states as possible, "because that is future"; but I think the idea was borrowed from a misinterpreted Biology.
In fact, a tertiary animal is more versatile (and bigger) than a secondary one. But the versatility is not much higher if the segments are too uniform. A colony, as some Tunicata/Hemichordata/Urochordata, is simply a cluster of many secondary animals superficially connected by a stolon or such. Even then, it helps to be safer from death, because if something partially eats it, the remaining segments restore the body.
Annelids are more integrated. The segments, obviously the secondary animals, are linearly arranged and the resulting tertiary is anterior-posteriorly ordered by Hoxes. But even the most homogeneous Annelida must have a single mouth and a single anus, so a few foremost and a few hindmost segments must differ. Four mouths and five anuses dispersed along the axis would seriously hinder the ordered operation of the alimentary channel.
Now, it seems that Hox genes are very good to define body parts linearly, along the anterior-posterior axis. And it seems that at least all segmented triploblastic animals have some sequence of Hox genes which seem to have common origin. Homology between Drosophila and mammal Hoxes are proven in many cases.
It seems that the arthropod set of Hoxes consists of c. 9 genes. The vertebrate sets seem to be composed of 13 (but wait a moment), in "more evolved" vertebrates with more than 1 sets, with minor differences [37]. In mammals & birds 4 parallel sets are identified (a-d), but none of them is complete, so finally we find 38 Hoxes. Then it is easy to understand the constancy of the borders of body regions in large groups of animals. The details can be found in App. E, but a Hox gene posterior in the sequence acts more or less more posteriorly along the body axis (the vertebral rod) too. And then it seems that the diversity of the body decreased in the transition to tetrapods, cca. at the border between Frasnian and Famennian.
Again, there is a minor problem with the words. The term "diversity" is suggested by the "biological diversity" which should be kept high according to Green slogans. However, either in English, or in Latin diversity/diversitas has a trace of "controversy" in itself, as a second meaning. There is a neutral, scientific, word, not implying a priori the controversies, close kin of the previous: variety/varietas. Indeed Medieval & Early Modern Latin used this word as a terminus technicus meaning that a region, country or continent had some multiplicity (of languages, fashions, religions &c.). So I will write "variety", but the decrease of biological variety will mean what a Green would mean as the decrease of biological diversity.
When somebody tries to defend the biological variety, that means first the gene pool. If the number of individuals of a species decreases, its variety decreases, genetic drifts & such may arise and at the end we may save the tiger but by chance only the purple-spotted diabetic tigers. Hovever for tertiary animals (as onyxophorans, insects, cephalochordata and us) it has another meaning as well: the vatriety of bodily regions. This is so because an annelid, arthropod or vertebrate consists a few dozen iterative subparts. The most of them may be uniform (as e.g. in annelids), but they may show a serious variety as well, as e.g. in insects, birds and mammals. As it is not too surprising, now we believe that the mechanism labelling the regions goes via the Hox genes; see App. E; for tetrapods the 4 parallel Hox sets enable much finer identification than a single one (as in Drosophila or Amphioxus). So an animal with 38 Hoxes may indeed express a great variety in its body; but it is not sure that it will do it.
In first approximation we may simply speak about the variety of the vertebral rod, because i) the soft body is unknown for fossils; and ii) the Hoxes order the body anterior-posteriorly anyway, so along the vertebral rod. In a mammal, the vertebral rod starts from the base of the skull (the head may be homologous to the apical end of the Amphioxus' chorda dorsalis, but can be an innovation "New Head" as well), and it contains the following regions going from head to tail:
cervical > thoracic > lumbar > sacral > caudal
The 38 Hoxes could identify even more regions, on the other hand the thoracic/lumbar border is not too definite at lots of reptiles. We may say that the cervic/thoracic border is the pectoral girdle, and the sacral region is the vertebrae where the pelvic girdle is attached.
In mammals the regions are clear enough even in fossils if only a part of the vertebral rod is preserved; in the different regions the forms of the individual vertebrae are different. The cervic region contains always 7 vertebrae (exc. Edentata) which bear no ribs (exc. Monotremata). The thoracic vertebrae. (c. a dozen) bear ribs, the lumbar, sacral and caudal ones do not; and the lumbar and sacral vertebrae are different for forms. As for the caudal part, it may be long or short, but anyways, the vertebrae are quite uniform except that they are narrower and narrower going endward.
However the last paragraph was not a tetrapod review, only a mammal one. The listed clearly different and specific vertebral regions are products of a long evolution. In primitive tetrapods often all or most vertebrae (including caudal ones) bear ribs or something so rib-like that I will call them ribs. And for the thoracic-lumbar border, in many cases that may be simply a mammal superstition.
Of course, the cervic and caudal ribs are generally shorter than the thoracic & lumbar ones; and the sacral ones must be either very short or absent for the proper working of the pelvis. You may consult [21] & [41]; the primitive tetrapod vertebral formula is well-developed long ribs in the whole thoracolumbar region, longest in midway. The clear distinction between thoracic and lumbar vertebrae started with the Cynodonta on the synapsid line and very probably was correlated with the appearance of the diaphragm at the thoracic/lumbar border (more explicitly: the place of that border is the diaphragm). In Procynosuchus the difference between the thoracic and lumbar ribs is recognisable, but only in details. In Thrinaxodon the lumbar ribs are still present, but the lenghts are very much reduced. Just before the mammals the tritylodontid Olygokiphus has only very rudimentary short cervic, lumbar and caudal ribs [41]. But this differentiation is a product of long evolution. Originally the pectoral girdle was simply at the base of the skull, the neck did not exist (just now the first fossil on the way to tetrapods with a neck is the fishapod Tiktaalik), and on the fish grade there were no recognisable thoracic, lumbar and caudal regions on the vertebral rod. The sacral region simply did not exist at all.
The explanation for the uniformity of the primitive vertebral rod is surely the lack of necessity for differentiation. The Hox sets can label regions, but there is no necessity of differences in the vertebrae. The Hoxes cause varieties mainly in the soft body and in the fins.
Now, let us see the status of the variety in the time of Eusthenopteron (Frasnian). Of course we know only the skeleton; but that we indeed know well. The vertebral rod is fairly uniform from the base of the skull until the beginning of the tailfin: the caudal ribs are longer than the more anterior ones. However Eusthenopteron is a sarcopterygian, so the fins have bony supports. Taking them into consideration too, we find the following regions (some terms are my suggestions, and even I am not fanatic about them, but I must call them somehow):
1) The head/skull. Maybe it never belonged to the vertebral rod; but in the linear sequence of Hox genes those characteristic for the head are on their proper places.
2) The neck/cervic region. Its length is still exactly 0 in Eusthenopteron (it will be 7 vertebrae in mammals).
3) The pectus/pectoral region. It is defined by the existing pectoral girdle, at the base of the skull. I cannot call it on the usual Latin name, because according to the anatomic consensus the humerus is the part of the limb/fin. Obviously long ribs would disturb the proper motion here in the joints.
4) The thorax/thoracic region. This is the region starting just behind the pectoral girdle.
5) The abdomen/lumbar region. The different names for the soft body and for the vertebral rod obviously come from anatomic conventions in the Rinascimento. The transition from the thorax cannot be seen at all on the vertebral rod of Eusthenopteron and surely it is gradual in the soft body; but after some more 150 Mys the region will be seen on the vertebral rod as the lumbar region.
6) The sacrum/sacral region. It cannot be seen in Eusthenopteron's vertebrae, but the skeleton virtually defines it as the intersection of the rod and a line between the bases of the bone to the anterior dorsal fin and the midpoint of the bases of the paired abdominal fins. This line may have been virtual (pointed out by the Hoxes, of course), but may also have been physically present as ligaments or cartilages.
7) The chironic region. This is the region between the sacrum and a similarly defined next point (see there); the name comes from the famous centaur Kheiron. As for the soft body, we are still anterior to the anus. Gonads certainly were there. (In us the region cannot be detected on the vertebral rod and is doubtful in the soft body, possibly about the gonads.)
8) The analis region. On the skeleton this is at the intersection of the vertebral rod and the line between the bases of the posterior dorsal and the anal fins. As for the soft body the anus is surely well defined by Hoxes, and on extant fishes the said point of the vertebral rod is near to the anus of the soft body. But in Eustenopteron this point is not merely virtual/unseen on the vertebral rod. The bases of the posterior dorsal and the anal fins bonily attach to the rod [32]. (In us these fins do not develop.)
9) The cauda/caudal region. It is more or less the region of the caudal fin, later tail; it starts just after the analis region and on the vertebral rod it is well defined by the end of vertebral uniformity: first the rib length increases.
See Fig. 1, from above and laterally. This is Eusthenopteronis varietas, the variety of the body regions of Eusthenopteron fordii in the Frasnian of Upper Devonian (except that the cervic region is still of zero length). It is richer/higher than the tetrapod variety, from Frasnian to present. It is not too different from that of recent Latimeria (and from that on the ordinary fishes, where, however, the fins are weaker, so their positions influence the body less). However I told how difficult for us simply to feel that our domestic animals/familiar partners have 5 limbs, not 4. Practically no human feels that our cousin Latimeria has 8 body regions, not 6. I am a physicist and a physicist can reach over human/mammal/tetrapod idiosynchrasies in the maximal extent; but sometimes also I forget about the chironic & caudal regions where we have only insignificant and maybe virtual rudiments. (Of course we have the analis region even if only in the soft body).
And now consider a centaur. It has not been really developed, because Eusthenopteron’s lineage stopped at the Frasnian/Famennian border (we do not yet know why). If it had not stopped, the trick of Ordo Pleuronectiformes may have had happened, i.e. a lateral wandering of one organ by c. 90°, in some tens of My. If so, then the anal & dorsal fins, bonily attached to the vertebral rod, may have had become optimal for locomotion. (Even until not, one can walk for a while with an unpaired hindmost leg; see Pearson’s Puppeteers in Niven’s stories.) And then, would have come terrestrial vertebral hexapodalism…
For any case, observe that the Greek Centaur Idea evolved into a bad direction. On centaurs of Classical times the second limb pair was homologous with horses’ forelimbs, and the third pair to horses’ hindlegs. So the pectoral girdle was repeated and the third limb pair was attached to the sacral region. This would have been evolutionally absurd even with Eusthenopteron unextinct.
However somewhat earlier than Classical times centaurs were believed not a human upper half on horse’s shoulders; see e.g. the “Athene temple” of Assos. Rather, take an erect human body, + a hindmost body part, with a hindmost limb pair, an anus & private parts and then a tail [1]. Now, such a being is conform with a surviving Eusthenopteron-like lobe-fin 367 My ago, and then a further evolution. Compare Figs. 2 & 1.
It is an important question, for which I do not yet have any answer, is that: why the great innovation of legs and walking was synchronous with the vanishing of the chironic region and 3 buds of fins. The understanding of this question will surely result in better understanding of the biological evolution in general. Until then I only can tell about my impression. In the Frasnian a few sarcopterigian species try to utilize the edges of the land masses; and they are successful. According to [31], they utilize the direct radiation of Sun; but that is indeed a source utilizable only when out of water. Then on the Frasnian-Famennian border "the sky falls on them", and a few million years later we see no more fishes trying to get out. As if fishes' "collective memory" or their superstition would have preserved the interdiction: "Thou shalt not leave the Water, else the Sky shall fall on thee and thy kin!" As for Acanthostega & Ichthyostega: they are already terrestrial. OK; they cannot yet procreate on land, but for any case their swimming ability is strongly reduced, just as the variety of their body regions has been reduced.
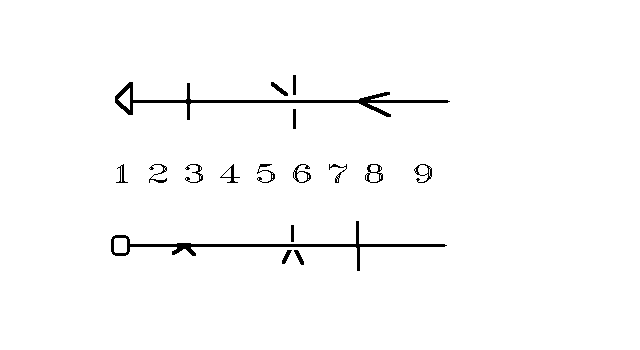
Fig. 1: The regions of the idealised Eusthenopteron-like transitional lobe-fin 367 My ago, from above and laterally. For Eusthenopteron Region 2 (the cervic one) was of 0 length, but the region already existed in that time for Tiktaalik roseae (which, however, rather belonged to Panderichthys’ lineage). For the definition of the regions see the text.
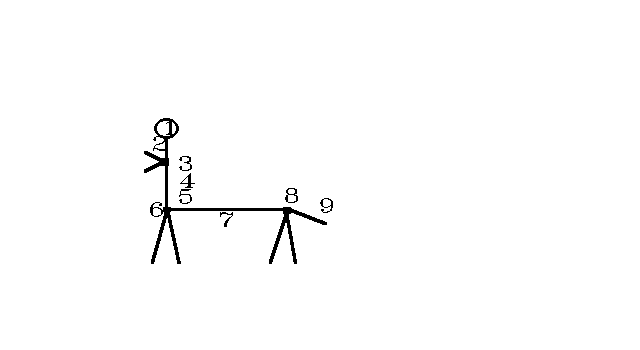
Fig. 2: The regions of a centaur, continuing Eusthenopteron’s lineage.
But of course if hexapodalism is not utterely impossible to be preserved during the conquest of lands, then surely the Galaxy should be full with hexapodal sophonts (Chap. 2). And then Fermi’s Paradox is even more serious…
And finally, AmphiHox set contains at least one gene more posterior than any in vertebrates. (Again, details are in App. E.) Maybe an anterior-posteriorly richer body plan, maybe hexapodalism was a virtual future for descendants of the LCA of Tunicata, Cephalochordata & Vertebrata, but it did not realize?
If so, then we may comment the Greek Centaur stories as: si non è vero, è ben trovato.
ACKNOWLEDGEMENT
The idea of the present study was provoked by the Janissary trilogy of Jerry Pournelle & al. In the story a small group of modern humans are hijacked & transplanted to a far planet, where, however, there is a human population originated from older transplantations; the newest happened during Charlemagne. The moderns start innovations including a University.
Now, the autochtones of the planet are centaurs, at an intelligence level somewhat above chimpanzees. What happens at the University when the moderns reach Darwin & Haeckel in Biology? Culture of Charlemagne was conform to God creating centaurs, but can that body plan be a product of “fishes” crawling to land?
My first, tentative answer is this study.
APPENDIX A: GREEK NATURAL SCIENTISTS AND THE CENTAURS
The first idea for the origin of us and the land animals which we can detect from later references is of Anaximander of Miletus. His idea was that the first men originated from (or within) fishes. (For an analogy see the vivipary of sharks; this seems to have been the mechanism reconstructed by Anaximander.) His argument was cca. that human babies need much more maternal care than newly-hatched fishes do; so "originally" humans were not able to live "in themselves" or "as they are now". The references seem rather unreliable to be more definite.
Empedocles of Akragas (495-435 BC) seems to have been used centaurs especially in his argumentation. His idea was that first not complete animals appeared but body parts. Then a centaur is possible; maybe they were not too successful. Aristotle refers him carefully, (see in due course), so we know more about him than about Anaximander.
Aristotle [42] mentions centaurs only once: at Bk 89b31-35 where he discusses logics. He discusses the question whether “a god or a centaur is or is not” in the sense if they do exist. The form of the question is piquant and interesting; and he does not answer this question. Obviously, his formulation suggests that a god or a centaur exist in not the same way, or not there as a horse or the City of Athens. He does mention chimeras as newborn with ram's head &c., but it seems he does not take them seriously. As for other monstrosities as newborn with more than two legs &c. he tries to give some explanations. In the Generation of Animals around Bekker page 723 he discusses Empedocles' "evolution theory" that first parts of animals appeared and combined freely. Such an idea would explain centaurs; but Aristotle is strongly against Empedocles' idea, because he states that such separate parts could not live.
Strabo, a few decades before Emperor Claudius, in his Geographika [43], writes a whole chapter about Arabia, but does not mention there any place called Saune and does not mention Centaurs in Arabia. He mentions Centaurs 4 times (VIII.3.19, IX.4.8, IX.5.12, IX.5.19) in connection with historical places in Greece; in the last case he comments them as savages, so there is no reason to believe them not human. About 180 AD Ptolemy in The Geography [44] lists many dozens of Arabian localities, none with a name even similar to Saune.
APPENDIX B: ON THIGHBONE FRACTURES IN THE UNIVERSE
It is possible to tell something about the maximal mass of bipeds in the Universe; and the result remains more or less true, with some caution, for tetrapods too. It will definitely not be a constraint for hexapods. In more details see [16].
Let us form the dimensionless combination of universal constants
αg
= Gmp2/hc = 6.2*10-39 (B.1)
where αg expresses the analogy to the fine structure constant
α ≡ e2/hc = 1/137.036
Then a man of 75 kg mass contains
Nh = αg-3/4 = 4.5*1028 (B.2)
nucleons. Observe that this number is the square root of the baryon number of an average star, or a neutron star,
N* ~ αg-3/2 = 2.0*1057
N* is almost exactly the maximal mass of a neutron star, the solar mass is 0.6N*, and the relation (B.2) is not an accident, as we shall see immediately, at least for order of magnitude. I give only some points of the proof.
A land animal cannot be so large that break when topples. Now, the binding forces are chemical, so essentially electromagnetic, while the breaking force comes from the action of gravity.
For simplicity, consider a cubic animal. Then
M ~ mpN(B.3a)
V = N1/3Va (B.3b)
Va ~ rB3 = (h2/mee2)3 (B.3c)
Only the third relation is not trivial; that is exact for a hydrogen atom, but now let us get it via the uncertainty relation
Δp
Δx ~ h/2 (B.4)
Since
Ek = p2/2me (B.5)
the minimal energy of the electron (i.e. when not moving) in the H atom at r average distance from the proton is (Δx=r/2)
E =
-e2/r + h2/2r2me (B.6)
and then the distance is just the Bohr radius. The binding energy is
EB
= e4/2h2 (B.7)
Now, a toppling cube gets a shock of energy MgV1/3/2, where g is the surface gravitational acceleration (g=981 cm/s2 for Earth), so the shock is ~N4/3. On the other hand, if it causes a break, it happens on an internal surface, so the energy is distributed amongst N2/3 atoms. So the energy/atoms goes with N2/3. This is the reason for an upper bound in N.
Substituting H atom data we would get M ~ 8000 g, too small by 1 order of magnitude. However matter with higher molecular weight is more compact. Anyways, we know that 75,000 g of (B.2) is almost exact, because thighbone fractures are sometimes lethal even now for older people while youngsters can topple without danger, so we ourselves must be near to the borderline. Humans are the biggest bipeds on Earth.
Tetrapods are safe if they step only with one leg in any moment. So elephants are safe until not running. Some padding, of course, helps. Hexapods would remain stable if stepping with not more than 3 legs, not on the same side. In the water uplift counteract weight (not inertia). So a whale is safe, until it does not hit something.
Now one can get a result valid anywhere in the Universe for land animals, except for so tricky lifeforms which we cannot yet even imagine. Namely, not every planet is able to develop life; some would not even support it for long time. We are now interested only in autochtones. You must then require that the planet
1) be at temperatures compatible with liquid water or ammonia for general solvent;
2) can keep its atmosphere, especially O2/CO2;
3) have a crust stable enough.
Now, the first condition means T=200-300 K. At such temperatures, as we experience it, Mars is already not massive enough to keep the atmosphere, so we cannot go too much below terrestrial mass because of Cond. 2. On the other hand, crusts are thinner with increasing planetary mass, so Cond. 3 does not permit planets significantly more massive than Earth, either.
Now, expressing everything via particle masses and fundamental constants we arrive at
N ~ [(me/mp)4/3α4(mpc2/T)]3/4αg-3/4 (B.8)
The prefactor is ~1 (I cannot yet tell if it is an accident or not), and the last term is just the "number magic" mentioned above.
I must confess that some dinosaurs were bipedal and still more massive than the upper border obtained here. Also, moas were some 3.5 m tall. Still, aging humans are often crippled or, after some weeks, die after toppling. See also [45].
APPENDIX C: ON TRIGLA
This Appendix has dual purpose: one personal and one important for the hexapod problem. First briefly the personal one.
In 1613, in Florence, a certain Lorini, a Dominican, made a mass which Galileo took as a personal attack. He was of course right: Lorini told something "Oh Galilean men, why you gaze into Heaven...", which was clear enough, and he declared that the teaching of a certain Pole Ipernicus contradicts Bible. Then Galileo started a counterattack. At a certain point Lorini told that i) he has tasks enough and this special problem is no great interest for him; ii) it is no matter if a Pole is called Ipernicus or Copernicus; and iii) he wanted to show that he also exists and can discuss the problem. So, let us see now the second purpose.
In Chap. 5 I mentioned a fish walking on the bottoms on the 3-3 digits of its pectoral fins, and told that it is the trigla or swallowfish, Exocoetus volitans. Now some readers can feel that I have made monkey of myself. But quiet; I know that something is not true here; only that is irrelevant in the possible evolution of pectoral fins, and in this way I got another possibility to demonstrate my knowledge.
Look: the taxonomy of fishes is still somewhat liquid. They are far from us, they live underwater &c. Then let us go step by step. There are 3 suborders relevant here, and it is even possible that they are only 2 [46], but let us continue. The suborders are Exocoetidea, Dactylopteroidea & Cottoidea. The textbooks identifying the trigla as E. volitans are good Hungarian textbooks (let me not tell the names of authors), but Hungary is land-locked and even before 1920 we had only a few kilometers of seashore (albeit just at the Adriatic). Trigla was fished by Croatians & Italians.
According to Ref. [46] (a precise German), Exocoetus volitans is Atlantic, while Dactylopterus volitans is Mediterranean. The latter fishes were called "swallows" in Antiquity, and Dactylopterus means "fin with fingers" seen on the trigla. So maybe not E. volitans but D. volitans is the trigla. However the situation is even more complicated: in the ample taxonomy of [46] we get
|
Subordo |
Family |
Genus & species |
Areal |
|
Exocoetoidea |
Exocoetidae |
Exocoetus volitans |
Atlantic |
|
Dactylopteroidea |
Dactylopteridae |
Dactylopterus volitans |
Mediterranean |
|
Cottoidea |
Triglidae |
Trigla gurnardus |
From Murmansk to Adria |
Now, the Triglidae walk on the "digits" of their pectoral fins. The Dactylopteridae also do this; and they "fly" too. The Exocoetidae are par excellence flying fishes, they glide with the pectoral fins, but have no "digits". Somebody may have confused 3 species of 3 suborders; the reason may have been the similarity of the latter 2, while the species names are the same for the first two. But of course, maybe fishermen of different Mediterranean shores simply called 2 different but outwardly similar fishes, D. volitans and T. gurnardus on the same name.
APPENDIX D: ON THE FIRST LAND ANCESTORS OF THE RECENT TETRAPODA
Centaurs (existent or not) call our attention to the point that i) if it were easy to create legs ex nihilo, then Vertebrata would have done it sometimes; and ii) if it is easy to attach fins/legs to the vertebral rod, then an explanation would be needed, why the third pair of fins of many fishes have been lost in us. It seems that it is not easy to form a third pair of legs ex nihilo (because of the Hox system?), but we do not yet know if the attachment of fins/legs was easy or not. If it was easy then it may have happened independently on different lineages. In principle we have lots of fossils plus the genetic information from extant tetrapods; maybe they will show if the appearance of the pelvis was a unique event or not.
Well, they will show it sometimes in the future. Just now the answer is equivocal. However let us see what can be known.
"Modern" (cladistic) and traditional (gradistic) terms for taxa sometimes contradict subtilely when using the same words, which fact disturbs the argumentation. [47] observes this, and gives a short glossary for the modern language. 3 taxon names are important for now:
Amniota: "the last common ancestor of mammals and reptiles, and all its descendants".
Amphibia: "extant lissamphibians and all extinct tetrapods that are more closely related to them than they are to amniotes".
Tetrapoda: "the last common ancestor of amniotes and lissamphibians, and all its descendants".
According to mathematical logics, everything is clear. If all present tetrapods are classified into 2 and only 2 groups in any definite way, then either both groups have their separate LCAs, or one group is ancestral to the other. But even in the second case the asymmetry in the definitions helps.
The cladistic language is clear, and in principle genetic analysis of extant members lead directly to the cladogram. However for fossils the information about kinship is indirect. Since for a long time the descent was based on fossils, let us see first the traditional system(s).
In the traditional (gradistic) system extant tetrapods belong to 4 Classes: Mammalia, Aves, Reptilia & Amphibia. (In cladistic language [extant] Mammalia, Aves & Reptilia form the extant part of Clade Amniota and extant members of Grade Amphibia belong to Clade Amphibia. The problem is with the extinct members.) Going "downward", Mammalia came from synapsid Reptilia (in cladistic language, Synapsida), the transition was so continuous that the "reptile-mammal" border is quite artificial. The mono- or diphylecy of the transition depends on this artificial boundary [48]. As for Aves, they came from amongst or besides the Dinosauria, so, depending on the system of "reptiles", they are modified Diapsida or "Triapsida". Now, the "reptiles" are obviously not monophyletic, but the Amniotes may be; or they may not. The presence of the amnion is a condition of completely terrestrial life; or thinking backwards, which is not irrational in evolutionary contexts, a consequence of it. Namely, in embryogenesis, the aqueous environment permits free-swimming larvae, but on land this is impossible. This is the present crucial difference between "amphibians" and "reptilians".
However even now there are amphibians which are not amphibian (note the 2 different meanings in one half-sentence) without aquatic swimming larvae (e.g. Salamandra atra); on the other hand, amniote (and even viviparous!) Ichthyosauria were fully aquatic, and Huehne [21] prefers that they were primarily aquatic; indeed their extremities do not show any trace of the tetrapod digit structure; they are clearly fins.
No doubt; as for procreation, amniotes are much better land-adapted than non-amniote tetrapods, but "Amniota" expresses the presence of an amnion, while "Amphibia" expresses only an amphibious lifestyle. And indeed, Huehne derived Ichthyosauria from Stegocephalia, as a line parallel to Anura. He might or might not be true. But note that in cladistic context the Amniota/Amphibia dichotomy is not more/less terrestrial or progressive/conservative at all. Sharks, ichthyosaurs, Latimeria and Vipera are all viviparous so for procreation "completely terrestrial". I guess that the first members of Clade Amniota did not yet have amnions, so the offspring started as aquatic larvae. Again, that name suggests something which it should not; remember Gorgias. The cladistic definition of Amniota is not a being with amnion, but exactly what was written above.
The monophylecy of the recent amniotes cannot yet be decided. Namely, the aortas of the amniotes develop in a dichotomic way: for recent snakes, lizards & birds the right aorta is more developed than the left one, while for mammals this is in the opposite way. Obviously the primitive status must have been symmetric, but we do not know how old is the dichotomy. For any case, Géczy [17] tells that the split may even have happened "on amphibian grade". Since an amphibian grade or lifestyle does not imply Clade "Amphibia" as opposed to Amniota, we are as we were, concerning the monophylecy.
Traditionally the classification of Reptilia went according to the extra openings of the skull, which are told to be irreversible. Apart from the 3 eye- and 2 nose-openings, the ancestral tetrapods had none. However in early reptile stage new openings started to develop. According to the positions and numbers the consensus was for a long time that 5 subclasses could be recognised:
|
Name |
N° of pairs of openings |
Position |
Skull |
|
Anapsida |
0 |
- |
Anapsid |
|
Parapsida |
1 |
above |
Parapsid |
|
Euryapsida |
1 |
above |
Euryapsid |
|
Synapsida |
1 |
middle |
Synapsid |
|
Diapsida |
2 |
above & below |
Diapsid |
Then turtles are Anapsida, we mammals are Synapsida, while snakes, lizards and birds are Diapsida; the other 2 branches are extinct: the ichthyosaurs were Parapsida and the plesiosaurs were Euryapsida.
So far so good. Still, Géczy suggested 10 (!) independent branches [17]: 2 instead of the old Anapsida, 2 instead of the old Euryapsida and 4 instead of the old Diapsida.
Géczy's narrower Anapsida subclass is simply the Ordo Cotylosauria, expiring in Upper Triassic; they were primarily anapsids. He is not sure about the ancestors of the other anapsids, the Chelonia/Chelonomorpha: according to him they may come from the Cotylosauria, or from Rhyncocephalia or from Plesiosauria, or, although it is not probable, even from Urodela. Now, in the latest case, Amniota would not be monophyletic. Since the "Amphibia" ancestry of the Ichthyosauria is rather seriously discussed as well (but not from Urodela [17], [21]), and the first synapsids (e.g. Protoclepsydrops) are synchronous with the first cotylosaurs, so surely different branches left the water in Carboniferous, I think it is obvious that monophylecy of Amniotes is not yet proven at all. Let us pass Amniotes now.
Present Amphibia form 3 orders: Apoda (Coecilia), Urodela & Anura. We know next to nothing about the ancestors of Apoda, and traditional but careful classifiers (see e.g. [21], [22]) were often in doubts about the monophylecy of the other two orders. I will return to this point, but the chaos is even greater. More than a decade ago I participated in demonstrating this chaos [49], and I do not think it has become clearer. Namely, when having included the extinct taxons of Classis Amphibia, the picture is as follows:
|
Devonian |
Ichthyostega |
|
Carbon. & Perm |
Lepospondyli & Labyrinthodontia |
|
Triassic |
Labyrithodontia |
|
From Jurassic |
Lissamphibia |
Labyrithodontia contain Ichthyostegalia, Temnospondyli & Anthracosauria, at least. (This is actually [53] .)
Now, the Ichthyostegalia are not the ancestors of the newer Labyrinthodontia, and even less of the Lepospondylii. The Lepospondyli die out completely in Permian, and the Labyrinthodontia seem to die out without descendents at the end of Triassic. On the other hand, the Lissamphibia appears in the Triassic, but without any visible connection to the last Labyrinthodonta.
Let us confront [17] & [21]. [17] lists 4 standpoints:
1) Porolepiformes → Urodela vs. Osteolepiformes → Anura.
2) Temnospondyli → Anura; Lepospondyli → Apoda; ?? → Urodela.
3) Anura & Urodela vs. Apoda.
4) Lissamphibii are monophyletic.
I think, it is easier to tell: we do not know. To be sure, [47] does not know either.
[21] agrees with Possibility 1). Let us note that two generations ago this scheme was popular, and it comes directly from Jarvik, who used an observation of Säve-Söderbergh, both Acanthostega/Ichthyostega experts [50], [51]. Jarvik started from the choana of osteolepiforms [51], reconstructed the same on porolepiforms (which is at best doubtful now; see e.g. [52] against it), and it started just turning out that the choana of the actinist Latimeria is not a true choana. Since the choana is important in breathing, the scheme was clear-cut: 3 related groups, Osteolepiformes, Porolepiformes & Actinista, the first 2 with choanae tend to go to be terrestrial, and the third without goes to deep sea. Urodela & Apoda are Pseudocentrophori (from the structure of the vertebrae); that group comes from Porolepidiformes and is extant. On the other hand, Anura comes from Osteolepiformes via Loxembolomeri (the family Loxommidae). So far so good; but there is a gap during all Permian.
And then we have a web of descent full of gaps. Ichthyostega and its many near relatives, the latter mainly found in the last decade, are good enough primitive Devonian tetrapods with the first traces of amphibious lifestyles; but the connection to Carboniferous known tetrapods is in doubt. (OK; if not Ichthyostega, then another Upper Devonian tetrapod was the ancestor; somebody will find it sometimes. Maybe Hynerpeton?) However then the well-known C-P Labyrinthodontia are not the ancestors of our Lissamphibia (some still unknown Labyrinthodonta may be; or may not), and Lepospondyli are no ancestors of anything seen after Palaeozoic.
And this is not surprising at all. Being amphibious is something living partly in water partly on land. Maybe in different ages different lineages had amphibious genera. And amphibious lifestyle leads to amphibious anatomy, in which case "convergent evolution" may mimic kinship.
Recently Anabas scandens or Periopthalmus koelreutheri are quite amphibious for lifestyle while they both are good Teleostei.
So: we do not know if the hindlegs were attached once to the (later sacral region of the) vertebral rod, or twice (an immediate precursor of Ichthyostega in Upper Devonian for Eutetrapoda and something similar to Adelogyrinus in the lowest Lower Carboniferous for Urodelida [21]) or even more than twice. Before amphibious times the vertebral rod did not show a recognisable sacral region at all, so the sacral region is the consequence of the hindlegs attached, therefore we identify a sacral region if hindlegs are attached. (Sorry for being repetitive.) Still if the trick had been repeated many times, the final results should have shown more differences than between the larval developments of Urodela & Anura.
Genetic trees might help, but the picture is not yet clear. [54] gives divergence times in a calibration where Synapsida/Diapsida split is 310 Ma. According to them Lissamphibia diverged from us at 360±15 Ma ago. So this is the "Amniota/Amphibia" split, coincident to the Devonian-Carboniferous boundary. (Sure, recent amphibians are all Lissamphibia per definitionem and we are amniotes.) However the standard error would permit both evolution of the sacral region once, in some immediate ancestor of Ichthyostega and only then an "Amniote/Amphibia" split and a split still before the attachment of hindlegs and independent terrestralisation on two or more lineages as well. [47] deals with this problem, so for details see it and citations therein.
[54] may support monophylecy of pelvis among recent Tetrapoda and for any case is against the Porolepidoformes origin of Urodela. Namely, as seen above, the Lissamphibia/Amniota split is 360±15 Ma, so between Upper Devonian and Lower Carboniferous. Now, the Porolepidoformes/Osteolepidoformes split is Lower Devonian or even older. If Urodela were Porolepidoformes descendants, one would expect larger standard error for the divergence time. Of course an Upper Devonian divergence for a diphyletic Lissamphibia is still possible if the two branches started from different Osteolepidoformes fishes. Candidates can be easily found.
And look: there is something strange about the Lissamphibia-Homo divergence time in [54]. The result 360±14.7 My is not the expectation value ± standard deviation; the 360 My is the mode. The mean is 441 My. 441 Mya would be Latest Ordovician. Since fans of The Avatar, readers of Greek mythology &c. find generally the details of the formalism of Mathematical Statistics boring, there will be an Appendix H, whither I relegate my notes on means, medians & modes.
Fig. 2i of [54] demonstrates that the distribution of the single-gene divergence times is very non-symmetric for this divergence. In itself this does not mean anything too definite, even if the single-gene distribution mimics the sum of two symmetric ones with means about c. 360 & 700 (!) Mys. However the strange distribution may be a signal of sampling error. Now, one sampling error may be an inhomogeneous Lissamphibia sample. If the present Lissamphibia is an artificial taxon, then almost anything can be obtained for divergence times. If Huehne & others were right then the mean of the divergence time would be something between the divergence time of Homo & Anura and that of the ancestors of Urodelidia & Eutetrapoda. This happened once in the not too far past when divergence times were calculated/estimated between Pongidae & Hominidae and they were in the range of 15-20 My; now we know that there is no such taxon which would contain Pongo, Gorilla & Pan but not Homo; and Pongo, the outlayer, increased the divergence time. In [54] the Homo-Pan divergence time is 5.5 My, and the ghost of the problem with Pongo & Sivapithecus still haunts somewhat there.
I think, something is still not understood about the circumstances at the Frasnian/Famennian boundary. A monophylecy of tetrapods, even that of amphibians, would mean that after Famennian the first tetrapods, already on land, repelled all the attempts of their near kins which were quite similar to them. And also, that in the next hundred millions of years all attempts made similarly to recent Anabas scandens or Periopthalmus koelreutheri were unsuccessful. This is possible: only not too probable and is still without any explanation. This monophylecy problem obviously is more complicated than the present study. If readers want to get a deeper insight to the problem, I can refer to the book [55] from 1991 (and especially the Chapters written by H. P. Schultze and J. R. Bott) and the article [56] from 1992. Of course, we are almost 2 decades later. I note only that in 1992 Carroll [56] was not yet sure about amphibian monophylecy.
And I do not go into details but also only note that the Urodela autopodium may be an innovation compared to all other known tetrapods [57], namely to all amniotes and Anura. This was just the standpoint of Huehne [21]; now the argument is about the expression of Hox-11 genes in the buds which are precursors of hands & feet, and the special formation of Urodela digits. True, [57] shows a solution keeping the monophylecy of Lissamphibia: a partial digit loss and then the de novo formation of at least 2 digits, sometimes in the Dark Ages of the Urodela. It is quite possible, of course, and this be enough for a while.
APPENDIX E: HOX GENES, AMPHIOXUS AND OUR LOST HINDMOST PART
As told in Chap. 9, Hox genes help to organise bilateral (segmented?) animals' bodies, along the body axis. But the limbs of tetrapods violate the linearity of the body. Are the fingers of the hand governed by forward Hoxes according to the origin point of the forelimb, or by some other Hoxes, or by no Hoxes at all?
Ref. [37] determines nicely the proper Hoxes for the origin of the limbs. The correspondence is conservative, from true fishes to us. The bud of the pectoral fins/arms forms in a region on whose posterior part the Hoxc-6 gene expression starts; obviously Hoxa-6 and Hoxb-6 makes this even more definite. (Hoxd-6 does not exist.) As for somites of early embryonic development, the bud region starts at Somite 10 in mouse, 17 in chick, 19 in goose, 3 in frog Xenopus and 2 in zebrafish; but the connection to Hoxc-6 is constant. The differences in somite numbers come mainly from the various numbers of cervic vertebrae. Mammals almost always have 7 cervic vertebrae, birds more but variable, and the zebrafish does not have any neck. As for the 7 mammal cervic vertebrae, Hoxd-4 & b-4 are expressed in all (the anterior border of the expressions is sharp for Hoxes), a-4 starts in the second, c-4 in the fourth, and Hox-5-s start in the sixth. So Hox-4's and -5's in themselves can explain almost all individual differences of the cervic vertebrae within the group [58]; then it is trivial to see the difference between cervic and thoracic ones.
The organisation of the vertebral rod into cervic, thoracic, lumbar, sacral & caudal parts is common in mammals & birds; surely it is common amniote development at least. (The sacral region being singled out by the hindlegs, e.g.)
Burke & al. mapped the expressions of Hoxes in mouse and chicken; only the anterior border is sharp, so the posterior one will not be mentioned. They got that Hox-4 & 5's are expressed in the cervic region, -6 - -9 are expressed in the thoracic one (but d-9 in the lumbar one so it seems that the distinction between thoracic & lumbar regions/vertebrae is younger than the fourfold multiplication of the Hox set); the sacral region is mainly c-10, d-10 and the 11's and the caudal region is the 12's & 13's. As for the boundaries, the cervic/thoracic border coincides with c-5/6, the thoracic/lumbar one (in mammals & therapsids the diaphragm) is defined in such a way that a-9, b-9 and c-9 are expressed at the posterior end of the thoracic region but all -10's are lumbar. Now, the lumbar/sacral boundary coincides with Hox-d9/10, but interestingly enough in chicken a-10 is lumbar while c-10 (and d-10) sacral. The sacral/caudal border seems to coincide with the beginning of Hoxd-11 activity.
Now, the 4 incomplete Hox sets in amniotes (and 7 in zebrafish) are obviously secondary via gene multiplications. It seems that in lamprey only 2 sets are present, and in amphioxus so far only a single set has been found. So maybe the amphioxus Hox set is the primitive state. Indeed, [59] reports that the amphioxus Hox set is conservative; but [59] also reports AmphiHox-14 & -15. Where are they expressed; and where are -12's & -13's expressed in highest primates without tails?
The answer for the second question will be somewhat surprising; the first question is somewhat ghastly in itself. It seems that Amphioxus has a so posterior part which we have lost; namely it has 2 Hox genes behind AmphiHox-13 [59], [60]. (To be sure, now we know that the vertebrate set contains a Hox-14 too, and it is reported from so close relatives of us as the coelacanth, but it seems lost in all tetrapods and all teleosts, at least. But Hox-15 is not reported so far from any vertebrate.) Anyways, because of more posterior Hox genes the Amphioxus may differentiate more the postanal regions (see e.g. [61]): then why does it not do it, or why do we not see the result?
As for the action of Hox-11, -12 & -13's, in land tetrapods it has been shown that they affect the hand & foot peripheries [62], mainly the digits! While this is contrary to the claimed collinearity, some genes must do it, and there is some analogy between most posterior and most distal. Also, one may perhaps contemplate about the ancestry of limb buds (but no paired anything on Amphioxus & lamprey). And Hox-14? And Hox-15?!
And now see the postsacral region of the vertebral rod. Note that in any extant tetrapod the tail is immediately after the sacral region.
Now, [37] compares Hox gene anterior expressions and starts of different vertebral regions between mouse & chick embryos according to somites. Of course both mouse and chick are tetrapodes.
|
Mouse somite |
Mouse Hox |
Region |
Chick Hox |
Chick somite |
|
5 |
*b-4 |
Cervic |
*b,d-4 |
5 |
|
19 |
c-6 |
Thoracic |
c-6 |
19 |
|
25 |
*a,b,c-9 |
Lumbar |
a,b,c-9 |
26 |
|
31 |
d-10 |
Sacral |
c,d-10 |
30 |
|
35 |
d-11 |
Caudal |
d-11 |
39 |
where “*” stands for "near to". The pattern is clear enough, similar enough, and there is no somite between the last of the future sacral region and the first of the future caudal one. There is one important region of the soft body at the sacral/caudal border or just behind: the anus.
However surely this was not so 370 My ago, and maybe not even now in Latimeria. It seems that Eusthenopteron still did not have a sacral region on its vertebral rod (at its abdominal fins, not attached bonily, the vertebrae are quite homogeneous); but the rod has a more posterior region where the anal & dorsal fins attach bonily. Only after that come the caudal vertebrae, different [31], [32] from the more anterior ones.
From chick & mouse one could guess that the region which was distinct in Eusthenopteron marked the anterior expressions of some -11's and maybe some -12's. The region must be the location of the anal & dorsal buds in Latimeria even if the buds are dead & lost in us forever. What can we learn about our lost vertebral region from Latimeria? Or have already something learnt which I do not know? [63] raised very apt questions 16 years ago. Anyways I am not ichthyologist or geneticist but a physicist from General Relativity & Heavy Ion Physics.
APPENDIX F: QUEER HYBRIDISATIONS ON ISLE OF MAN?
D. I. Williamson, marine biologist of the Port Erin Marine Laboratory, disturbs the orthodox biologists since 1988 by reporting transphylan hybrids. To be sure, they are not superstitious rumours but scientific reports in scientific journals. The idea is written down in a book published at Kluwer Academic [64], which is regarded as a solid scientific editorial house. The most detailed reports are about hybrids of a male Echinus esculentus (a sea-urchin) and a female Ascidia mentula (an urochordate, or by another taxonomy, a tunicate). Since Echinodermata and Tunicata are disjoint (even if neighbour) phyla, such hybridisations seem to contradict to XXth century genetics, XIXth century biological common sense, antic sceptics as Lucretius Carus, and everything we learnt to esteem. So the usual reaction is contaminated sperm, the androgynous nature of the sea-urchin and such.
Williamson's opinion is that he, of course, was very, very careful; the hybridisation is real; but it is not exactly as we see e.g. in mules. It is rather in connection with the strange larvae.
Larval stages of ontogeny are sometimes similar to the adult even if they are not identical. The well-known examples are e.g. the tadpole of frogs and the axolotl. The tadpole of frog is completely aqueous, for a time it has only hindlegs. Then transforms (or finishes the hatching), and we have the frog. The axolotl is the larval stage of a normal salamander (which, however, does not exist in nature but can be produced in laboratory). It has external gills, its legs are very weak, and it is fully aqueous. In Late XIXth century following Haeckel biologists told that Ontogeny Recapitulates Phylogeny; the tadpole & axolotl show the earlier, pre-amphibious Past. Now we are not so sure about the slogan, but even then, the ontogeny starts from a single cell, so the early stages cannot be minuscule copies of the adult. And in the tadpole & axolotl we see the homologies of most organs with those of the adult; if the gills and for the frog the tail are completely lost, that is more or less trivial.
However in many cases the larva's Body Plan is quite different from that of the adult. The well-known example is the butterfly whose larva is a caterpillar. However this is not the most surprising example. The evolutional tree suggests roughly onychophoran ancestors for insects, and if Ontogeny Recapitulates Phylogeny, then...
However medusae & polyps of cnidarians differ much more in Body Plan; similarly some Echinodermata have very, very surprising larvae, and in some cases the larva of the deuterostomate echinoderm definitely seems protostomian!
Williamson believes that in the rare transphylan hybridisation he sees the recapitulation of the process when two animals "made an alliance" for life. More or less one animal became the young phase of the other. He believes that so distant "hybridisations" mean the adding up of the genetic matter. The result is generally not viable (too different genes &c.), but with proper regulation the time periods of the workings of the incompatible genetics will not overlap, and then we saw the very strange larvae.
This may and may not true. He believes that the "acquisition of larvae" was successful only in the times of Great Crises of Life as the Devonian, Permian and end-Cretan mass extinctions. If the successful hybridisation is very, very rare but the crisis kills off all non-allied members of two distant species...
His opinion is that his successful hybridisation was simply the reiteration of earlier evolution; so in the very rare cases when the offspring reached maturity, it started as the larva of one parent, then the larva hatched and the other parent emerged. We cannot combine sea-urchin and urochordate characters. In first approach even Nature, with unnumbered experiments and hundreds of million years cannot; the two beings are simply put "one after another". (In second approximation some genes may wander between larva and adult, and then...)
It is interesting that the idea of alliance of two very distant species as different lifestages because of crisis of life appears in the sci-fi of Orson Scott Card, Xenocide, which is a sequel to Ender's Game. It seems that the first publication of the book was 1991, while Williamson's book Larvae and Evolution was published only in 1992. Of course, journal publications existed since 1988.
APPENDIX G: ON THE ORIGIN OF THE CENTAUR IDEA
Since the centaur idea was persistent in Classical Greece, something must provoked it. The simplest explanation is a popular one that some non-riding Greeks misinterpreted non-Greek horsemen.
If this is the origin, the time must be the Early Dark Ages not much after the fall of the Mycenian civilisation. Namely, Mycenian warriors were not riding; they were charioteers. On the other hand, while Homer knows this, his description is utterly erroneous about charioteer way of fighting. So chariots went out of use and then some, possibly foreign, elite appeared on bigger horses, unnaturally sitting on them. Only: Homer’s reports speak about savages, not an elite.
APPENDIX H: MEANS, MEDIANS & MODES
We see in [54] that the divergence time for the Homo-Lissamphibia split, calculated from 95 genes, is 360±15 My if we ask for mode; but in expectation value (mean) it is 441 My. Slightly strange; and then what is the age of the split: Famennian or Lower Silurian? That is really a difference. If the mode gives the correct information, then Lissamphibia is monophyletic and diverged from our lineage already in the wet sand and Jarvik & Huehne were wrong; but if the mean is informative then even all Lissamphibia may come from porolepiforms.
I cannot tell what happened in the wet Famennian surfs; for example because there is some peculiarity with the X axis of Fig. 2i of [54]. However let us see the definitions of mean, median & mode. That will help a little. [65] is a rather formal reference; the definitions are the same in any textbooks.
Consider first a finite set of numbers {xi} (say, you measured a quantity n=2m+1 times). Then you order the numbers to be monotonously increasing, and then
|
Mean |
<x> = ∑xi /n |
|
Median |
xC = xm |
So the median is just the middle value. As for the mode, you count, how many times the same x value was obtained. The mode is the value obtained most times. If that is not unique, the distribution is polymodal.
In physics we generally act as if the number of measurements were >>1; anyways it would be good to have a lot of result, and in many times we could repeat and repeat the measurement (if needs be). So we like to think in continuous probability distributions
f = f(x) (H.1)
where, by definition
∫f(x)dx = 1 (H.2)
since the sum of the probabilities of all imaginable results is of course 1. If we perform n>>1 measurements, we can somehow approximate the true distribution f(x) (e.g. via interpolations).
Now, the mean, or expectation value is
<x> = ∫xf(x)dx (H.3)
As for the median, you first calculate the integral probability F(x)
F(x) = ∫-∞xf(x')dx' (H.4)
and for the median xC:
F(xC) = 0.5 (H.5)
the central one. Finally, the mode is the most frequent value, so the place of the maximum of f:
f(xmode) = max{f} (H.6)
If the distribution f(x) is symmetric to a value of the x variable x0, i.e.
f(x0+y) = f(x0-y) for all y (H.7)
and f is homogeneously increasing until x0, then x0 is simultaneously the mean, the mode and the median, as follows from (H.3-6). Otherwise they generally do not coincide.
Which is the truest one? In general the question is ill-defined. If, however, we measure a definite quantity and we know what resulted in the distribution, sometimes something can be guessed. In [54] the quantity was the divergence time T, which was calculated from different genes. Everyone gave an approximation ti, and then [54] accepted the mode (H.6).
What comes is a toy model, whose simplifying conditions generally do not hold. However, because of averaging away, central distribution theorem & such it is generally not a bad idealisation.
Take a quantity which is the average of several independent quantities (so they fluctuate independently):
x = ∑ xI/N
around, say, 0, with mean deviations σI. If N→∞, then the resulting distribution goes to the limiting distribution which is Gaussian
f(x)→(1/2πσ2)exp{-x2/2σ2} (H.8)
where
σ2 = ∑σI2/N (H.9)
Such more or less independent additions come from noises. So if a time value t is determined to be something, but with a number of disturbing factors, then in first approximation we expect
t = T + x (H.10)
with the above distribution for x. Of course, it is not sure that the noises are additive. However, e.g. for multiplicative noises
1) the Gaussian distribution is simply replaced by the log-normal one; and
2) for small noises the multiplication can be linearized and we are back at (H.7).
So if the divergence time has a more or less symmetric (moreover: more or less Gaussian) distribution, then probably everything is almost perfect. If not, then at least one assumption does not hold.
Now, the distributions of Figs. 2a-h of [54] are more or less symmetric, therefore the means and modes almost coincide, so we do not have to choose. But not on Fig. 2i, the lissamphibian-human divergence time. I, of course, do not know what happened there. I mentioned one possibility: that the Lissamphibia are diphyletic, so the sample is inhomogeneous. Indeed, then one expects a distribution with either 2 peaks at the two real divergence time, or with a plateau between the 2 values. However, there is a simpler possibility.
Assume that the "noises" appear in a quantity y directly connected with mutation probabilities & such. Then, of course, there would be a Gaussian distribution centered at y=Y. But we cannot measure y. However if
y = at + b (H.11)
(uniform molecular clock), then the distribution is Gaussian in t too, and everything is OK.
However, let us go beyond linearity. if
y = y(t) (H.12)
then
f(t) = exp{-(y(t)-Y)2/2Σ2}(dy/dt)-1/(2πΣ2)
y = y(t) (H.13)
Then the distribution will not be Gaussian, and generally not even symmetric, for 2 reasons:
1) There is a distortion in the exponent; and
2) there is a multiplicative function in (H.13).
E.g. assume that the mutation rate (or at least the rate of surviving mutations) was low between t1 & t2. The bins used were equidistant in t. However then the bins are unequal in y, in which the distribution is expected to be roughly Gaussian. Between y1=y(t1) and y2=y(t2) the distribution seems to be too low, because the molecular clock ticked there slowly.
I repeat, I do not know what happened in the Lissamphibia-Homo split time calculation. I only know that if the mean and the mode are too different, then a physicist would not take the mode. Namely the detailed form of the distribution is generally more sensitive to errors than the expectation value (which is an integral over all values so the majority of errors is expected to "average out"), or the median, which is calculated from the integrated probability (the same argument).
And if we have any doubt in the real equidistancy of bins, the mode will be directly sensitive on the equidistancy.
The end-Devonian is far enough in the past to be not sure about the uniform ticking of the molecular clock. The mean would be less sensitive; but it would give 441 My; not error, that is small, but perhaps bias. In addition if the lissamphibian sample is inhomogeneous, i.e. not monophyletic, then the divergence time may be simply an artefact. If the mean & the mode were near to each other, then that would be a self-consistency argument for monophylecy, but they are not even similar to each other...
REFERENCES
[1] D. E. Strong: A Lady Centaur. Brit. Mus. Quart. XXX, 36 (1965)
[2] Apollodoros: Bibliotheke, ed. J. Frazer. Loeb Classical Library, London, 1954
[3] R. Graves: The Greek Myths. Penguin, Harmondsworth, 1955
[4] J. L. Borges, Margarita Guerrero & N. Th. di Giovanni: The Book of Imaginary Beings. Penguin, Harmondsworth, 1980
[5] Plutarch: Seven Sapientium Convivium. In: Moralia, Vol. 2. Loeb Classical Library, 1928
[6] Pliny: The Natural History, Book 7, Ch. 3. H. G. Bohn, London, 1855
[7] Phlegon of Tralles: Book of Marvels, ed. W. Hansen, University of Exeter Press, 1996, Chaps. 34-35
[8] Lucretius Carus: On the Nature of Things, ed. W. E. Leonard, 1916, Book V
[9] J. Bergström: Opabinia and Anomalocaris, Unique Cambrian Arthropods. Lethaia 19, 241 (1986)
[10] S. J. Gould: Wonderful Life. Hutchington Radius, 1989
[11] S. Conway Morris: The Crucible of Creation: The Burgess Shale and the Rise of Animals. Oxford University Press, Oxford, 1998
[12] A. Seilacher: Vendobionta and Psammocorallia: Lost Constructions of Precambrian Evolution. J. Geol. Soc. London 149, 607 (1992)
[13] A. Seilacher: A. Fossil Art. Royal Tyrrel Museum of Paleontology, Alberta, 1997
[14] T. P. Crimes & D. McIlroy: A Biota of Ediacaran Aspect from Lower Cambrian Strata on the Digermul Peninsula, Arctic Norway. Geol. Mag. 136, 633 (1999)
[15] Sz. Bérczi & B. Lukács: Symmetry Versus Katachi in Animal Organisms. Forma 16, 91 (2001)
[16] B. Lukács & Gy. Paál: A világ szerkezeti állandói. Csillagászati Évkönyv 1982, Gondolat, Budapest 1981, p. 250
[17] Géczy B.: Ősállattan, Tankönyvkiadó, Budapest, 1979
[18] M. I. Coates: The Devonian tetrapod Acanthostega gunnari Jarvik: Postcranial anatomy, basal tetrapod interrelationships, and patterns of skeletal evolution. Transact. Roy. Soc. Edinburg: Earth Sci. 87, 363 (1996)
[19] J.-J. Hublin: L'évolution de vie. Flammarion, 1981.
[20] M. I. Coates & J. A. Clack: Polydactily in the Earliest Known Tetrapod Limbs. Nature 347, 66 (1990)
[21] F. R. Freiherr von Huehne: Paläontologie und Phylogenie der Niederen Tetrapoden. VEB Gustav Fischer, Jena, 1956
[22] A. S. Severcov: Vvedeniye v teoriyu ëvolyucii. Izdatel’stvo moskovskogo universiteta, Moscow, 1981
[23] F. H. Collins: Epitome of the Synthetic Philosophy. D. Appleton & Co, New York, 1895
[24] Lynn Margulis, M. F. Dolan & R. Guerrero: The Chimeric Eukaryote: Origin of the Nucleus from the Karyomastigont in Amitochondriate Protists. PNAS 97, 6954 (2000)
[25] M. J. Chapman: One Hundred Years of Centrioles: the Henneguy-Lenhossek Theory, Meeting Report. Internatl. Microbiol. 1, 233 (1998)
[26] E. N. Marais: Die Siel van die Mier. Johannesburg, 1925.
[27] A. Desmond: Archetypes and Ancestors: Paleontology in Victorian London, 1850-1875. Chicago University Press, Chicago, 1984
[28] B. Hatschek: The Amphioxus and Its Development. MacMillan, New York, 1893
[29] A. Willey: Amphioxus and the Ancestry of the Vertebrates. MacMillan, London, 1894
[30] Aristotle: Complete Works, ed. J. Barnes, Princeton University Press, Princeton, 1995. Bekker N° 94a24-b27 (Posterior Analysis)
[31] R. L. Carroll, J. Irwin & D. M. Green: Thermal Physiology and the Origin of Terrestriality in Vertebrates. Zool. J. Linnean Soc. 143, 345 (2005)
[32] S. M. Andrews & T. S. Westoll: The Poscranial Skeleton of Eusthenopteron foordi Whiteaves. Transact. Roy. Soc. Edinburgh 68, 207 (1970)
[33] P. Claeys, J-G. Casier & S. V. Margolis: Microtektites and Mass Extinctions: Evidences for a Late Devonian Asteroid Impact. Science 257, 1102 (1992)
[34] P. Claeys & J-G. Casier: Microtektite-like Impact Glass Associated with the Frasnian-Famennian Boundary Mass Extinction. Earth Planet. Sci. Lett. 122, 303 (1994)
[35] K. Wang: Glassy Microspherules (Microtektites) from an Upper Devonian Limestone. Science 256, 1547 (1992)
[36] Nóra Fáy, B. Lukács, K. Martinás & Sz. Bérczi: On Cosmic Spherule Composition: Gravitation+Thermodynamics. Internatl. Symp. Planet. Impact Events & Their Conseq. on Earth, PIECE '99, ed. Y. Miura, Yamaguchi, 1999, p. 25
[37] Ann C. Burke & al.: Hox Genes and the Evolution of Vertebrate Axial Morphology. Development 121, 333 (1995)
[38] P. E. Ahlberg: Postcranial Stem Tetrapod Remains from the Devonian of Scat Craig, Morayshire, Scotland. Zool. J. Linnean Soc. 122, 99 (1998).
[39] A. Sedwick: On the Origin of Metameric Segmentation and Some Other Moirphological Questions. Quart. J. Micro. Sc. XXIV, 43 (1884)
[40] B. Lukács: Vertebrata: Individual or Colony? Symmetry 4, 139 (1993)
[41] T. S. Kemp: Mammal-like Reptiles and the Origin of Mammals. Academic Press, New York, 1982
[42] Aristotle: Complete Works, ed. J. Barnes, Princeton University Press, Princeton, 1995
[43] Strabo: Geography. Loeb Classical Library, 1924
[44] C. Ptolemy: The Geography. Dover, New York, 1991
[45] B. J. Carr & M. J. Rees: The Anthropic Principle and the Structure of the Physical World. Nature 278, 605 (1979)
[46] K. Deckert & al.: Urania Tierreich. Urania Verlag, Lepzig, 1966
[47] M. Laurin, M. Girondot & A. de Ricqlès: Early Tetrapod Evolution. Trends in Ecol. and Evol. 15, 327 (2000)
[48] Sz. Bérczi & al.: Monotremes: About the Role of the Permotriassic and Jura-Cretaceous Boundaries in the Atherian-Therian Divergence. In: Proc. Symp. „Side Tracks in Evolution“, eds. Sz. Bérczi & al., KFKI-1998-07, p. 20
[49] Sz. Bérczi & B. Lukács: Main Lines and Side Tracks: The Highway of Amphibians Seems Wide Enough. In: Proc. Symp. „Side Tracks in Evolution“, eds. Sz. Bérczi & al., KFKI-1998-07, p. 13
[50] G. Säve-Söderbergh: Notes on the Dermal Bones of the Head in Osteolepis macrolepidotus. Ag. Zool. Bidr. Uppsala 20, 523 (1941)
[51] E. Jarvik: On the Structure of the Snout of Crossopterygians and Lower Gnathostomes in General. Zool. Bidr. Uppsala 21, 237 (1942)
[52] G. Clement: Evidence for Lack of Choanae in the Porolepiformes. J. Vert. Paleont. 21, 795 (2001)
[53] E. Kuhn-Schnyder & H. Rieber: Paläozoologie: Morphologie und Systematik der ausgestorbenen Tiere, G. Thieme, Stuttgart, 1984
[54] S. Kumar & S. Blair Hedges: A Molecular Timescale for Vertebrate Evolution. Nature 392, 917 (1998)
[55] H. P. Schultze & Linda Trueb (eds.): Origins of the Higher Groups of Tetrapods: Contoversy and Consensus. Cornell University Press, Ithaca, 1991
[56] R. L. Carroll: The Primary Radiation of Terrestrial Vertebrates. Ann. Rev. Earth Planet. Sci. 20, 45 (1992)
[57] G. P. Wagner & al.: Evolution of Hoxa-11 Expression in Amphibians: Is the Urodele Autopodium an Innovation? Amer. Zool. 39, 686 (1999)
[58] G. S. B. Horan & al.: Compound Mutants for the Paralogous Hoxa-4, b-4 and d-4 Genes Show More Complete Homeotic Transformations and a Dose-Dependent Increase in the Number of Vertebrae Transformed. Genes & Development 9, 1667 (1995)
[59] Linda Z. Holland & al.: The Amphioxus Genome Illuminates Vertebrate Origins and Cephalochordate Biology. Genome Res. 18, 1100 (2008)
[60] T. P. Powers & C. T. Amemiya: Evidence for a Hox-14 Paralog Group in Vertebrates. Curr. Biol. 14, R183 (2004)
[61] D. E. K. Ferrier & al.: The Amphioxus Hox Cluster: Deuterostome Posterior Flexibility and Hox-14. Evolution and Development 2, 284 (2000)
[62] J. Zákány & al.: Regulation of Number and Size of Digits by Posterior Hox Genes: A Dose-dependent Mechanism with Potential Evolutionary Implications. PNAS 94, 13695 (1997)
[63] M. I. Coates: The Origin of Vertebrate Limbs. Development 120, S169 (1994)
[64] D. I. Williamson: The Origin of Larvae. Kluwer Academic, Dordrecht, 2003
[65] H. Cramer: The Elementary Probability Theory. Wiley, New York, 1955