Originally published in Symmetry 4, 161 (1993)
VERTEBRATE: INDIVIDUAL OR COLONY?
B. Lukács
Central Research Institute for Physics, H-1525 Bp. 114. Pf. 49., Budapest, Hungary
ABSTRACT
Serious problems about the finer details of the internal deuterostomic structure of complicated topology raise the question if the vertebrates are secondary or tertiary animals, meaning the steps in which they were built up from living constituents. This question is discussed in the paper.
1. INTRODUCTION
Not too successful individuals or peoples often look for successful aliens as kinfolk or ancestors. However, in Regnum Animalia we Vertebratae are the most successful ones (according to our common opinion) and still our own ancestors and kinfolk are not too popular objects of study; other Deuterostomatae are rather neglected as compared to such popular Protostomatae as Annelidae, Insectae or Cephalopodae. Before continuing this Introduction, Fig. 1 shows a very schematic deuterostomid half of an evolutionary tree, and Fig. 2 gives pictures of characteristic members of the deuterostomid phyla, after Ábrahám (Összehasonlító állatszervezettan, Budapest, 1964), Dudich Állatrendszertan, Budapest, 1967) and Géczy (Őslénytan, Budapest, 1981).
|
|
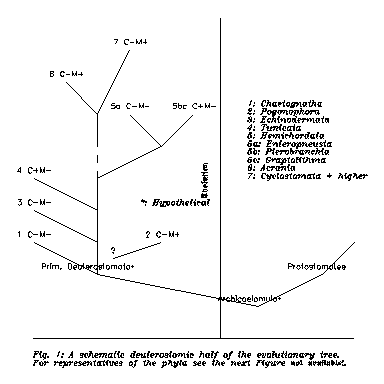
Fig. 2: The original Fig. 2 never existed in digital form. It gave sketches of: Sagitta hexaptera, Stylophora, Appendicularia sicula, Glossobalanus minutus, Rhabdopleura Normani & Amphioxus.
The Deuterostoma family album starts with Sagitta hexaptera for Chaetognata. The picture does not show yet anything resembling a chorda dorsalis, and it is difficult to see if the neural network is ancestral to a spinal cord. For any case it starts from the neighbourhood of the mouth, but it is natural to get a neural center at the apex.
Our second kin is not recent. It is Stylophora, mentioned earlier [1] for its expressed lack of a bilateral symmetry. Recent discussions suggest that it was not an Echinodermata proper but rather something between positions 3 and 4. In this case the opening at the apex is a mouth, the opposite one is the anus, the tail may have contained a chorda dorsalis, and the thickened part beside the anus housed a brain.
The next picture shows Appendicularia sicula, belonging to Tunicata. There is a well developed chorda dorsalis in the tail, and parallelly on its dorsal side runs a neural fiber analogous to the spinal cord.
Glossobalanus minutus represents position 5a. The e is a so called notochord in the glans at the apical end, not seen here, it is sometimes regarded as the analogon of the chorda dorsalis. Rhabdopleura Normani stands for 5b. These individuals form a colony connected via the stolon; in each member from the mouth two channels start, the enteral one (through the stomach to the anus) and the stomochord (which may be the analogon of the dorsal chord).
Finally Amphioxus (Branchiostoma lanceolatum) stands for Vertebratae. No need to bore the reader here with so familiar further kinfolk as Cyclostomata, Pisces, &c., showing no important difference from us at all. Amphioxus will be discussed later in some details, so now observe only the segmented muscles, the many repeated gills, the homogeneous spinal cord and the lack of any mentionable head at the apical end. (The head will develop in later stages of evolution.)
Let us start with a problem in connection with the statement of an earlier lecture [1]. There it was demonstrated that deuterostomatae are complicated for topology, and this complicated structure may have given the possibility for higher complexity, effectivity or anything else behind our feeling of superiority. Thus far this is only a hypothesis, not discussed further. However one may ask about the steps leading to the more complicated topology, and then it turns out that in this point the picture is still partially obscure. This may indicate a fairly complicated evolution, with some very special steps.
Let us mention only one possible "anomaly". If one tries to visualize the possible steps to reverse the rôles of mouth and anus, then the simplest reconstruction is the one mentioned in Ref. 1: a primitive Coelomata had distorted itself into U-shape, then the intestinal channel worked improperly at the turning point, so there a secondary channel developed with a new opening, and this new one has become a mouth instead of the opposite, for any reason of its own. The functionless half of the channel gave a possibility to house something new and useful: either the spinal or the dorsal cord. Then the branching point of the enteral and neural channels (or that of the first and the chorda dorsalis) is expected at the neighbourhood of the mouth.
It seems to be so for Chaetognata (intestinal and neural channels) and for Hemichordata (intestinal channel and stomochord). However, the embryonal development of Amphioxus (Acrania) suggests a different story. The very schematic picture is Fig. 3 (after Hatschek and Ábrahám): compare this with the corresponding Figure of Ref. 1 and the difference is obvious. Now the bifurcation point is at the anal end of the mature organism. We do not have to accept Haeckel's biogenetic law to see the problem: cases of human spina bifidia are conform with the structure observed at Amphioxus.
Fig. 3: The original Fig. 3 never existed in digital form. Amphioxus embryo in the stage of 5 stomites (after Ábraham & Hatschek) (I advise to look for Ábrahám (Összehasonlító állatszervezettan, Budapest, 1964), and for B. Hatschek (Studien über die Entwicklung des Amphioxus. A. Hölder, Wien, 1881).) Observe the developing U-shape of the "enteral channel". The dorsal branch will later house the spinal cord. Legends: A: protostoma; B: canalis neuroentericus (the branching point); C: neuroporus (closing later). Consider the points disturbing any simple evolutionary scheme: i) the canalis neuroentericus is at the protostoma, so will be at the anal end of the mature animal (if it is deuterostomic), compare this with Rhabdopleura on Fig. 2; ii) the development process of the neural channel seen here cannot reflect phylogeny, because then for many generations the animal could not eat; iii) the dorsal branch of the channel here does not contain endoderm at all, so does not seem a part of the original enteral channel.
So something special may have happened on the Vertebrata branch of the evolution, between the recent Amphioxus (Acrania) and the common ancestor of Acrania and Hemichordata or Tunicata or both, anyway well in the Precambrian. We do not know this common ancestor from fossils just as any ancestor of Acrania is unknown, so there is still room for speculation. (We would rather avoid fruitless discussions about the exact location of Acrania on the evolutionary tree; the term Vertebrata will be used in a broad sense including Acrania. Anyway, Amphioxus has a well developed chorda dorsalis, and with enough imaginative power one can bridge the gap between Acrania and Cyclostomata.)
Here we want to discuss the possibility that one critical step creting "anomalies" may have been a unification. More specially: in this paper we discuss some arguments for an evoluti-onary path leading from a colony of primitive Deuterostomatae (visualizable as analogons of Enteropneusta) to an integrated organism whose parts started thereafter to specialise. We are well aware that the idea is not new, was suggested a century ago by Hatschek [2], but was opposed by such relevant people as Haeckel [3] and Spencer [4]. Still, no harm in rediscussing the possibility after a century.
2. POSSIBLE CLASSES FOR COMPOSITE ORGANISMS
All Metazoa have, of course, living parts, similar to some independent living organisms. E.g. Parazoa consist of some cells similar to recent Flagellatae. Then, going beyond the statement that something is composite, it is useful to describe the relations between the animal studied and its at least virtually living parts. A lot of classifications may be relevant; in this Chapter we discuss three.
The first and oldest one is based on the existence of parts and subparts. Then an animal may be of [4]
1) Primary, having no separable parts. Examples: Protozoa.
2) Secondary, having separable primaries. Examples: lower Metazoa, as Platyhelminthes; Mollusca.
3) Tertiary, having separable secondaries. Example: Annelida, whose segments more or less resemble individual lower vermins.
A further, quartiary step would be the integration of tertiaries into a single organism. A candidate is an anthill of very integrated insects, as was philosophically suggested by Marais for termites [5]. However in this classification one cannot decide if the organism is indeed a unit or not. But remember this example for further use.
Let us see our place in this classification. The classical authors [3,4] agree that we are secondaries. One may recapitulate the arguments following Spencer:
The Vertebrata (just as Mollusca) organism does not show any tendency to spontaneously separate into parts in any stage of ontogeny.
No homologous segments appear; the apparent segments are not original but come from adaptation (e.g. to undulating swimming); the vertebrate animal cannot be divided into segments containing most of the vital organs (as digestive, respiratory and reproductive ones for example). The Vertebrata is a secondary organism superficially segmented by external influences (while the segmented insects are true tertiaries).
Therefore the only secondary organisms showing up large and complicated forms would be molluscs and vertebrates.
In this Chapter we list, not judge, possibilities. So now let us go to the second possible classification, according to the degree of integration and autonomy. For details see Ref. 6 and further citations therein. We use 3 degrees:
A) Confederation. Mainly independent parts with interactions. Examples: sponges (separated cells survive but for a different kind of life, until reestablishing the secondary organism); an earthworm (can be cut into two and parts may live for a while to regrow the lost parts).
B) Federation. Identifiable separate parts, but with some vital functions located at the level of the whole organism. A separated part can survive for a while but with restricted functions; it cannot regrow the whole organism and cannot live its independent life. Examples: an insect (can survive decapitation for a short time, but cannot become a complete insect anymore); an anthill of termites (from which a termite can be separated for a short time but will die without reproduction &c.).
C) Unity. No possibility for survival of the separate parts for any mentionable time. Example: majority of higher animals (whose amputated fingers do not survive at all).
Obviously we belong to Case C): an amputated human limb cannot survive as an individual organism in any sense.
Now we introduce a third viewpoint into the classification, according to the origins of the parts. Several logical possibilities may exist but it is useful and conform with European traditions to think in triads. So here we define two extremal cases and every other transitional ones will be classified simply between them:
a) Symbiosis of originally independent animals.
ß) Transitional cases.
g) Self-generation of new segments.
Here it is difficult to give examples because we should know the prehistory of the composite organism. However, for Case a) one can mention the Eucariotes of endosimbionta origin [7], while Case g) is suggested by the ontogeny of the tapeworm or the Pogonophora (ancestry unknown). We will return to Case ß) after having some suggestions to specify.
Obviously the 3 classifications above (for composition, autonomy and origin) are not fully independent; not all the 27 combinations are possible, e.g. if something has no parts, then it must be a unity too. Again, one would expect higher autonomy for originally independent parts than for self-generated ones, but this is only an expectation and may depend on the history of the animal as well.
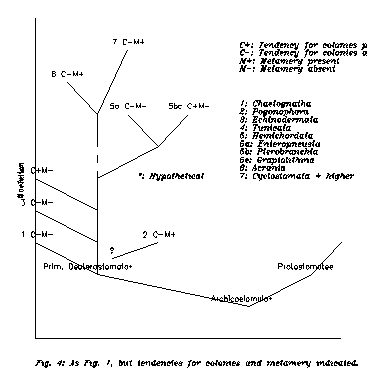
We close this Chapter with Fig. 4, which is a completed version of Fig. 1. The conclusions from this Figure will be postponed till the second Chapter from here.
|
|
3. ON AMPHIOXUS
The author is no expert of Amphioxus, this Chapter is no part of an Amphioxus monography, only a list of some facts not quite conform with Spencer's firm opinion (above) that no vertebrate could be divided into homologous segments carrying most of vital organs. But first we would like to emphasize that Vertebratae possess a 600 My evolution independently of any other phylum (say Hemichordata or Tunicata) and this time is quite enough to obliterate traces of autonomy by integration or those of homology by dissimilation, as seen e.g. on Metannelidae. Also, our favourite Amphioxus is not an ancestor of other recent Vertebratae, up from Cyclostomatae: its origin is quite obscure, and we can only hope that it may be a "living fossil". Still, with sufficient imaginative power one can bridge the gap between Acrania and Cyclostomata. Now let us see some vital organs or functions of Amphioxus:
Respiration: Gills, with 80-100 openings.
Circulation: No heart; pulsating sections of an artery, as many thickenings as the number of gills.
Secretion: As many protonephridia as gills.
Reproduction: 22-25 pairs of gonads in segmental arrangement.
Sensors: A single olfactory cavity in forward position (possibly defunct). Sensitivity to chemical stimuli on the whole surface although enhanced at the apex. Optical sensitivity in diffuse cells of the spinal cord, segmentally arranged.
Neural system: Central part: spinal chord in segmental arrangement + giant fibers connecting the segments. Dilatation at the apical end ("brain"). Dominantly gray matter for local functioning. Periphery: segmentally arranged neurons to muscles.
The above data definitely do not suggest a primordially unsegmented animal whose body became secondarily segmented for easier undulating motion. Such an adaptation might lead to segmented backbone muscles, but segmentally repeated eyes or gonads are not needed for easy swimming. Rather, the picture suggests a primordial homologous metamery, say between the homology of recent earthworm and insect, both always recognised unambigously as good tertiary animals.
4. ON THE POSSIBLE ORIGIN OF METAMERY
Now let us return to Fig. 4, which gives some suggestions. Lower phyla are generally oligomeric. The exception is Pogonophora. Then the first possible idea is that on the deuterostomic branch the tendency to grow multiple parts appeared independently, namely at the stage of Pogonophora, and this tendency is manifested in Vertebratae, but above Acrania in a rather obliterated way. To multiply the body is a very economic way (from informatical viewpoint) to be larger and more versatile, and such a body is a useful startpoint for later evolution, so if the tendency has appeared, it will probably be preserved by selection. In this case our position in the 3x3 classification is 3Cγ.
However in this way we run at least into 2 difficulties:
1) The tendency to self-generate segments seem absent in some higher units as Echinodermata and Enteropneusta.
2) In Pogonophora the metameric part is the opisthosome at the very end of the body. This part does not seem homologous to the segmented middle parts of Vertebratae, or to any part of them.
Fortunately one does not have to be disturbed by the above difficulties because recent rRNA sequence analyses have classified Pogonophora among Protostomatae [8]. Since in the lack of enteral channel the ventro-dorsal discrimination is ambigous in this phylum, we may accept the result. Having removed Pogonophora, Fig. 4 suggests the following pattern. Lowest Deuterostomatae are solitary and not metameric in the same time. First appears the tendency for forming colonies by cloning. Note that both for Pterobranchia and for Tunicata the slightly connected individuals are secondary unsegmented animals. The phylum Hemichordata may represent the transition: some classes have the tendency of colony-forming, some do not. Then going higher this tendency disappears, but "in its place" metamery can be observed. This pattern gets a natural explanation if the segmented body is the inheritor of a colony, and the individual segments are inheritors of the individual members of the colony. Here it is worthwhile to note that in the Cambrian colonial Graptolithinae were quite common. While this time is slightly late for direct connection to ancestral Vertebratae, it is a signal for a strong colony-forming tendency in the close neighbourhood of our ancestors.
We must confess that the present argumentation lacks constructive suggestions for the way of verifying or disproving the conjecture. Amphioxus' ancestors are not seen in fossils, and its embryogenesis does not seem to give a clear key [1], which is not surprising after the adaptations of 600 Mys. Haeckel's Biogenetic Law cannot be taken in face value on this timescale. Still, we would like to recommend the above picture as a viewpoint when looking at ourselves. In what follows we do not question the picture but draw the logical conclusions.
5. CONCLUSIONS
If the proto-vertebrates were analogous to a colony of Graptolithinae, Pterobranciae or Tunicatae, with later integration and dissimilation, then
1) In the 3x3 classification our place is 3Cß, where ß stands for "colony of semi-independent clones". This is indeed a transitional case between "symbiosis of aliens" and "self-generation of new segments", since the symbiotic individuals are of common genetic origin (identical twins or clones), but they are not limbs or segments but almost independent individuals (connected only by stolons). The ancestral colony would be 3Aß, the more integrated stage of the proto-vertebrate is maybe 3Bß. The actual place of Amphioxus could be measured by the extent of its regenerative power.
2) The specific parts of the present vertebrate body are not homologous to those of Chaetognata and Enteropneusta. So any "anomaly" may be expected when comparing the latter ones to Amphioxus.
3) The segmental arrangement of Amphioxus' gonads gets a natural explanation as remainder of the original state. Indeed, in this case there is no serious genetic presure to eliminate the multiple sources of reproduction, since the genetic information is the same in each segment (except for the rare mozaicism) they being clones.
4) Our classification 3Cß generates disturbing philosophical ideas concerning ourselves. They will be given now at the end of the paper.
We feel ourselves indivisible units. However this feeling in itself has nothing with our origin or even with the constitution of our body. Our self-consciousness is generated simply by the neocortex of the brain, which is not repeated in the body. However, looking at Amphioxus, one sees that the original vertebrate central neural system is the spinal cord. This organ is segmented, and its apical part specializes later into a brain. The Amphioxus seems to have preserved the very first steps of this specialization with a "brain" not qualitatively different from the posterior sections. Since a segment of the spinal cord possesses its own "eyes" and chemical sensors, and the intersegmental connections are not very strong, it is possible that the Amphioxus has "multiple personality just as the human society has: each segment can analyse data and command its own muscles independently, however with interaction and harmonisation between neighbouring personalities and with some "leading" ones at the apex. It is possible, of course, that contemporary Amphioxus is already past of this stage: maybe this point could be checked via teaching a definite segment by conditional reflexes and then looking for answers in the neighbours.
Is it so then that our original multiple personality is simply suppressed by the dominant mind(s) at the apex (now head), via the superabundant white matter conducting intersegmental commands, so that we cannot feel anymore the original multiple personality, and still the specialisation has been making the usurping apical segments so efficient that after some 600 Mys they can find out something about the original situation and can pay the obligatory respect to our own posterior, suppressed, atrophied and finally neglected personalities?
ACKNOWLEDGEMENT
The author would like to thank illuminating discussions with I. Molnár, who, however, is not responsible in any extent for the above speculative picture.
REFERENCES
[1] Sz. Bérczi, B. Lukács & I. Molnár: this Volume, p. 123
[2] M. Kretzoi in: Az élővilág fejlődéstörténete, ed. Tasnádi Kubacska M., Gondolat, Budapest, 1964 (in Hungarian)
[3] E. Haeckel: Über die Individualität des Thierkörpers. Jenaische Zeitschrift für Naturwissenschaft, XII, 1878
[4] F. H. Collins: A Summary of Herbert Spencer`s Synthetic Philosophy.5th Edition, London, 1901
[5] E. N. Marais: Die Siel van die Mier. Johannesburg, s.d.
[6] B. Lukács: this Volume, p. 19
[7] T. Cavalier-Smith: Ann. N. Y. Acad. Sci. 503, 17 (1987)
[8] J. A. Lake: Proc. Natl. Acad. Sci. USA 87, 763 (1990)
My HomePage, with some other studies, if you are curious.